
Response of Key Metabolites during a UV-A Exposure Time-Series in the Cyanobacterium Chlorogloeopsis fritschii PCC 6912


Abstract
:1. Introduction
2. Materials and Methods
2.1. Organism and Growth Conditions
2.2. Experimental Design
2.2.1. Supplemented UV-A (UV-A + PAR) Exposure
2.2.2. Time-Series Sampling
2.2.3. GC–MS Analysis of Intracellular Metabolites
2.2.4. Carotenoid Analysis
3. Results
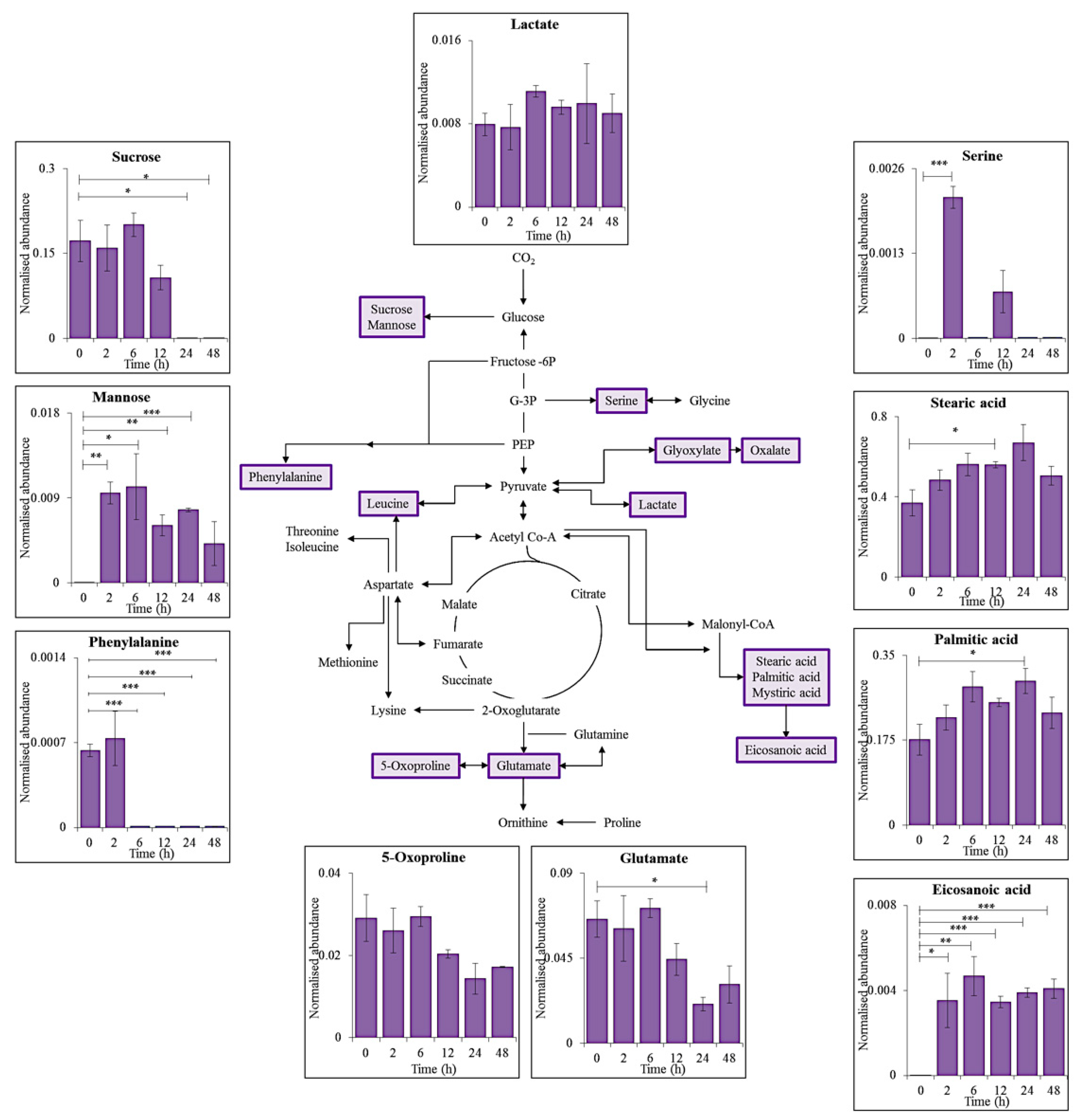
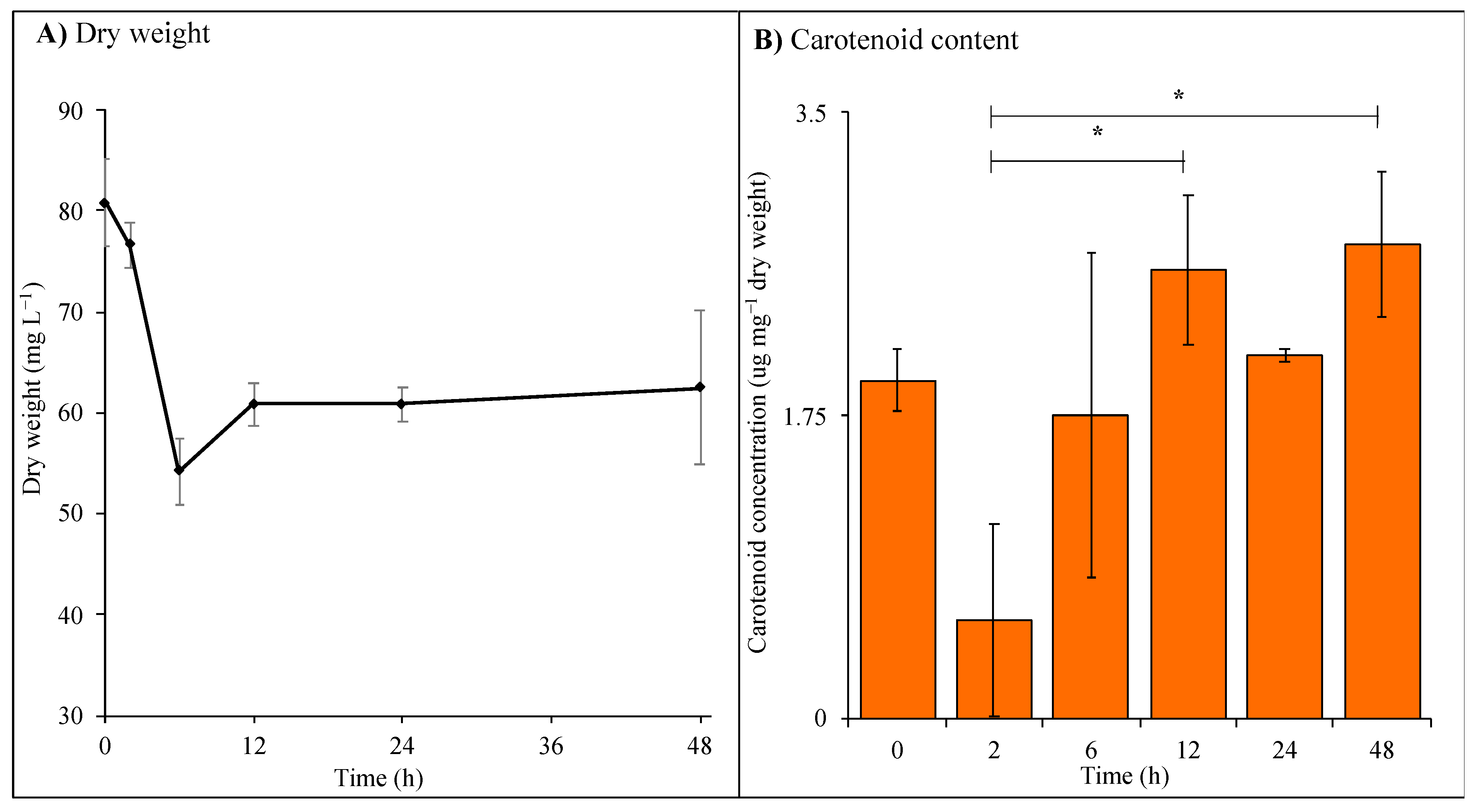
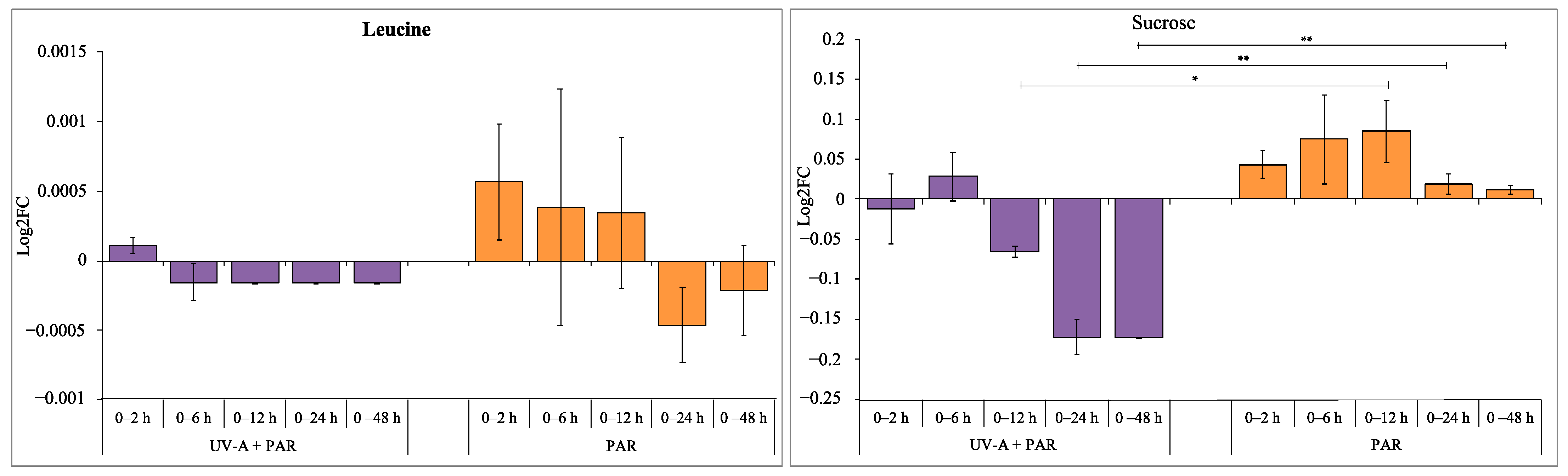
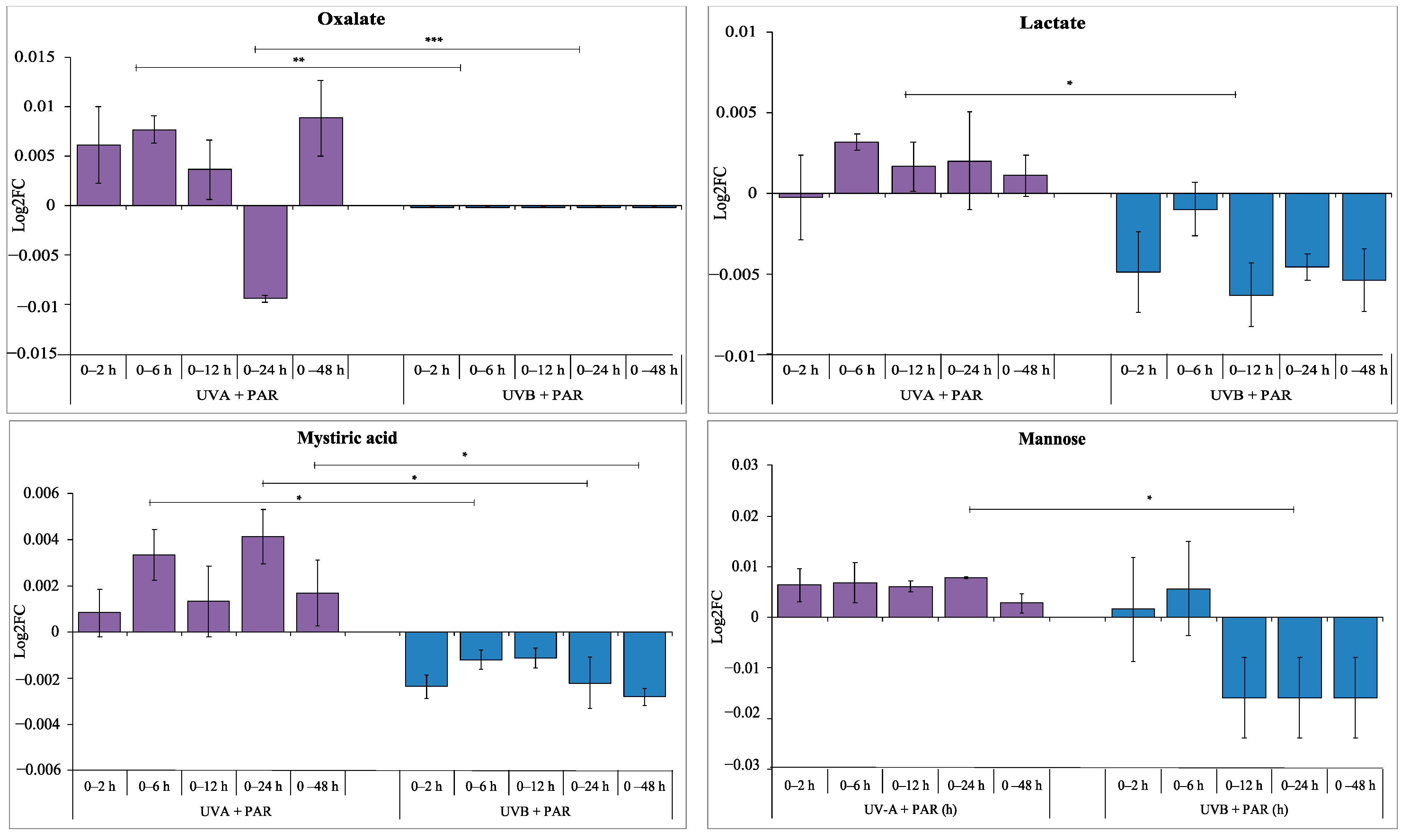
3.1. Intracellular Metabolite Levels during UV-A Exposure
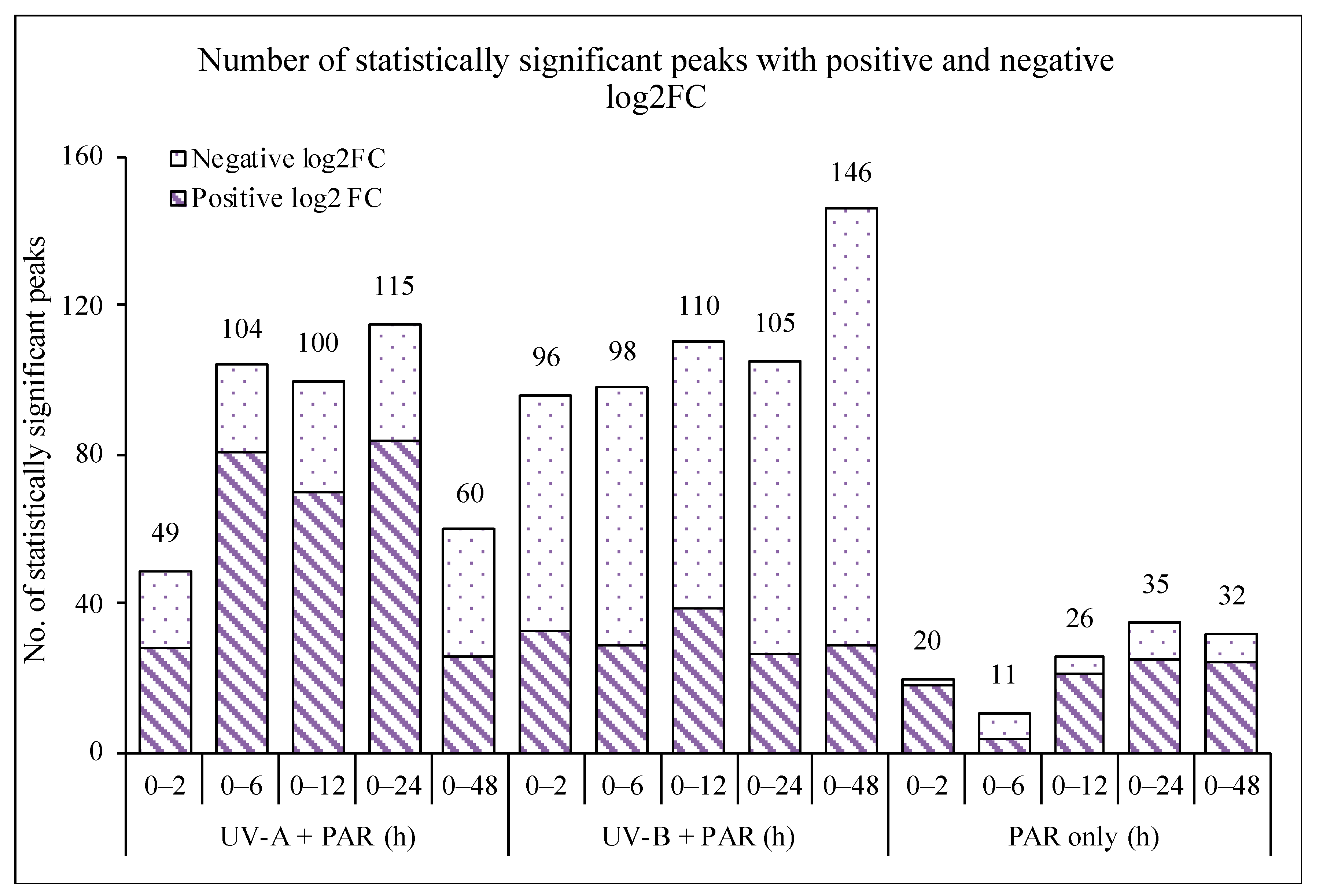
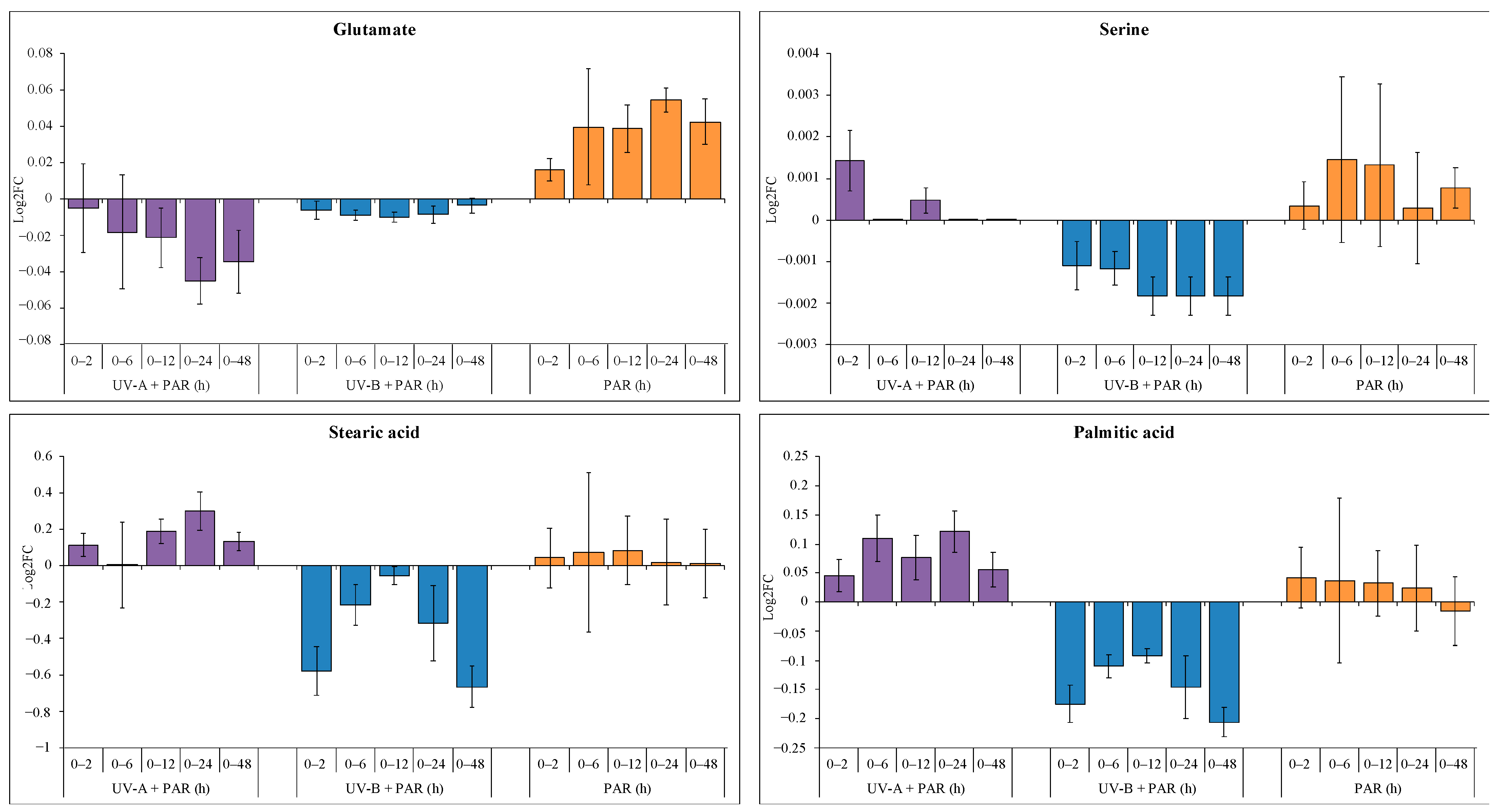
3.2. Comparison of Supplemented UV-A, UV-B and PAR Only
4. Discussion
Effects of UV Exposure on C. fritschii
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castenholz, R.W.; Garcia-Pichel, F. Cyanobacterial Responses to UV Radiation. In Ecology of Cyanobacteria II; Springer: Dordrecht, The Netherlands, 2012; Volume 9789400738, pp. 481–499. ISBN 9789400738553. [Google Scholar]
- Sinha, R.P.; Häder, D.P. UV-protectants in cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P.; Moh, S.H.; Lee, T.K.; Kottuparambil, S.; Kim, Y.J.; Rhee, J.S.; Choi, E.M.; Brown, M.T.; Häder, D.P.; et al. Ultraviolet radiation and cyanobacteria. J. Photochem. Photobiol. B Biol. 2014, 141, 154–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, Y.-J.; Kim, S., Il; Chung, Y.-H. Sensing and Responding to UV-A in Cyanobacteria. Int. J. Mol. Sci. 2012, 13, 16303–16332. [Google Scholar] [CrossRef]
- Sinha, R.P. Biochemical characterization of sunscreening mycosporine-like amino acids from two Nostoc species inhabiting diverse habitats. Protoplasma 2014, 252, 199–208. [Google Scholar] [CrossRef]
- Latifi, A.; Ruiz, M.; Zhang, C.C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.; Duarte, C.M.; Agustí, S. Contrasting Responses of Marine and Freshwater Photosynthetic Organisms to UVB Radiation: A Meta-Analysis. Front. Mar. Sci. 2017, 4, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Pascual, J.; Cañal, M.J.; Escandón, M.; Meijón, M.; Weckwerth, W.; Valledor, L. Integrated Physiological, Proteomic, and Metabolomic Analysis of Ultra Violet (UV) Stress Responses and Adaptation Mechanisms in Pinus radiata. Mol. Cell. Proteom. 2017, 16, 485–501. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Gao, K.; Villafañe, V.E.; Watanabe, T.; Helbling, E.W. Effects of solar UV radiation on morphology and photosynthesis of filamentous cyanobacterium Arthrospira platensis. Appl. Environ. Microbiol. 2005, 71, 5004–5013. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Scherer, S. UV protection in cyanobacteria. Eur. J. Phycol. 1999, 34, 329–338. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sinha, R.P. Biotechnological and industrial significance of cyanobacterial secondary metabolites. Biotechnol. Adv. 2009, 27, 521–539. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, M.; Bajguz, A.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Uncovering potential applications of cyanobacteria and algal metabolites in biology, agriculture and medicine: Current status and future prospects. Front. Microbiol. 2017, 8, 1–37. [Google Scholar] [CrossRef] [Green Version]
- Portwich, A.; Garcia-Pichel, F. Ultraviolet and osmotic stresses induce and regulate the synthesis of mycosporines in the cyanobacterium Chlorogloeopsis PCC 6912. Arch. Microbiol. 1999, 172, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Kultschar, B.; Dudley, E.; Wilson, S.; Llewellyn, C.A. Intracellular and Extracellular Metabolites from the Cyanobacterium Chlorogloeopsis fritschii, PCC 6912, During 48 Hours of UV-B Exposure. Metabolites 2019, 9, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D.; Incharoensakdi, A. Characterization and antioxidant functions of mycosporine-like amino acids in the cyanobacterium Nostoc sp. R76DM. Algal Res. 2016, 16, 110–118. [Google Scholar] [CrossRef]
- Joshi, D.; Mohandass, C.; Dhale, M. Effect of UV-B Radiation and Desiccation Stress on Photoprotective Compounds Accumulation in Marine Leptolyngbya sp. Appl. Biochem. Biotechnol. 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Klisch, M.; Walter Helbling, E.; Häder, D.P. Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation. J. Photochem. Photobiol. B Biol. 2001, 60, 129–135. [Google Scholar] [CrossRef]
- Shen, S.G.; Jia, S.R.; Yan, R.R.; Wu, Y.K.; Wang, H.Y.; Lin, Y.H.; Zhao, D.X.; Tan, Z.L.; Lv, H.X.; Han, P.P. The physiological responses of terrestrial cyanobacterium: Nostoc flagelliforme to different intensities of ultraviolet-B radiation. RSC Adv. 2018, 8, 21065–21074. [Google Scholar] [CrossRef] [Green Version]
- Wase, N.; Pham, T.K.; Ow, S.Y.; Wright, P.C. Quantitative analysis of UV-A shock and short term stress using iTRAQ, pseudo selective reaction monitoring (pSRM) and GC-MS based metabolite analysis of the cyanobacterium Nostoc punctiforme ATCC 29133. J. Proteom. 2014, 109, 332–355. [Google Scholar] [CrossRef]
- Schwarz, D.; Orf, I.; Kopka, J.; Hagemann, M. Recent Applications of Metabolomics Toward Cyanobacteria. Metabolites 2013, 3, 72–100. [Google Scholar] [CrossRef] [Green Version]
- Abed, R.M.M.; Dobretsov, S.; Sudesh, K. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef]
- Styczynski, M.P.; Moxley, J.F.; Tong, L.V.; Walther, J.L.; Jensen, K.L.; Stephanopoulos, G.N. Systematic Identification of Conserved Metabolites in GC/MS Data for Metabolomics and Biomarker Discovery. Anal. Chem. 2007, 79, 966–973. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. In Current Protocols in Molecular Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; Volume 131, pp. 30.4.1–30.4.32. ISBN 1552-4825. [Google Scholar]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. [email protected]: The Golm metabolome database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, J.; Sinelnikov, I.V.; Wishart, D.S. MetATT: A web-based metabolomics tool for analyzing time-series and two-factor datasets. Bioinformatics 2011, 27, 2455–2456. [Google Scholar] [CrossRef]
- Ritchie, R.J. Consistent Sets of Spectrophotometric Chlorophyll Equations for Acetone, Methanol and Ethanol Solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Henriques, M.; Silva, A.; Rocha, J. Extraction and quantification of pigments from a marine microalga: A simple and reproducible method. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. 2007, 2, 586–593. [Google Scholar]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, A.; Albert, A.; Ganzera, M. Effects of elevated ultraviolet radiation on primary metabolites in selected alpine algae and cyanobacteria. J. Photochem. Photobiol. B Biol. 2015, 149, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Cameron, J.C.; Pakrasi, H.B. Essential Role of Glutathione in Acclimation to Environmental and Redox Perturbations in the Cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2010, 154, 1672–1685. [Google Scholar] [CrossRef] [Green Version]
- Kolman, M.; Nishi, C.; Perez-Cenci, M.; Salerno, G. Sucrose in Cyanobacteria: From a Salt-Response Molecule to Play a Key Role in Nitrogen Fixation. Life 2015, 5, 102–126. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UV-A Exposure (h) | No. of Peaks, 12p < 0.05 a | No. of Peaks, Positive log2FC | No. of Peaks, Negative log2FC |
|---|---|---|---|
| 0 vs. 2 | 49 | 28 | 21 |
| 0 vs. 6 | 104 | 81 | 23 |
| 0 vs. 12 | 100 | 70 | 30 |
| 0 vs. 24 | 115 | 84 | 31 |
| 0 vs. 48 | 60 | 26 | 34 |
| PAR Only (h) | No. of Peaks, p < 0.05 a | No. of Peaks, Positive log2FC | No. of Peaks, Negative log2FC |
|---|---|---|---|
| 0 vs. 2 | 20 | 18 | 2 |
| 0 vs. 6 | 11 | 4 | 7 |
| 0 vs. 12 | 26 | 21 | 5 |
| 0 vs. 24 | 35 | 25 | 10 |
| 0 vs. 48 | 32 | 24 | 8 |
| UV-B Exposure (h) | No. of Peaks, p < 0.05 a | No. of Peaks, Positive log2FC | No. of Peaks, Negative log2FC |
|---|---|---|---|
| 0 vs. 2 | 96 | 33 | 63 |
| 0 vs. 6 | 98 | 29 | 69 |
| 0 vs. 12 | 110 | 39 | 71 |
| 0 vs. 24 | 105 | 27 | 78 |
| 0 vs. 48 | 146 | 29 | 117 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kultschar, B.; Dudley, E.; Wilson, S.; Llewellyn, C.A. Response of Key Metabolites during a UV-A Exposure Time-Series in the Cyanobacterium Chlorogloeopsis fritschii PCC 6912. Microorganisms 2021, 9, 910. https://doi.org/10.3390/microorganisms9050910
Kultschar B, Dudley E, Wilson S, Llewellyn CA. Response of Key Metabolites during a UV-A Exposure Time-Series in the Cyanobacterium Chlorogloeopsis fritschii PCC 6912. Microorganisms. 2021; 9(5):910. https://doi.org/10.3390/microorganisms9050910
Chicago/Turabian StyleKultschar, Bethan, Ed Dudley, Steve Wilson, and Carole Anne Llewellyn. 2021. "Response of Key Metabolites during a UV-A Exposure Time-Series in the Cyanobacterium Chlorogloeopsis fritschii PCC 6912" Microorganisms 9, no. 5: 910. https://doi.org/10.3390/microorganisms9050910