
Microalgae Cultivation on Nutrient Rich Digestate: The Importance of Strain and Digestate Tailoring under PH Control

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Digestates
2.2. Experimental Design
2.2.1. Culture Conditions and Inoculation
2.2.2. pH Manipulation
2.3. Growth Measurements
2.4. Nutrient Analysis
2.5. Determination of Total Proteins and Carbohydrates
2.6. Determination of Fatty Acid Methyl Esters (FAMEs)
2.7. Statistical Analysis
3. Results
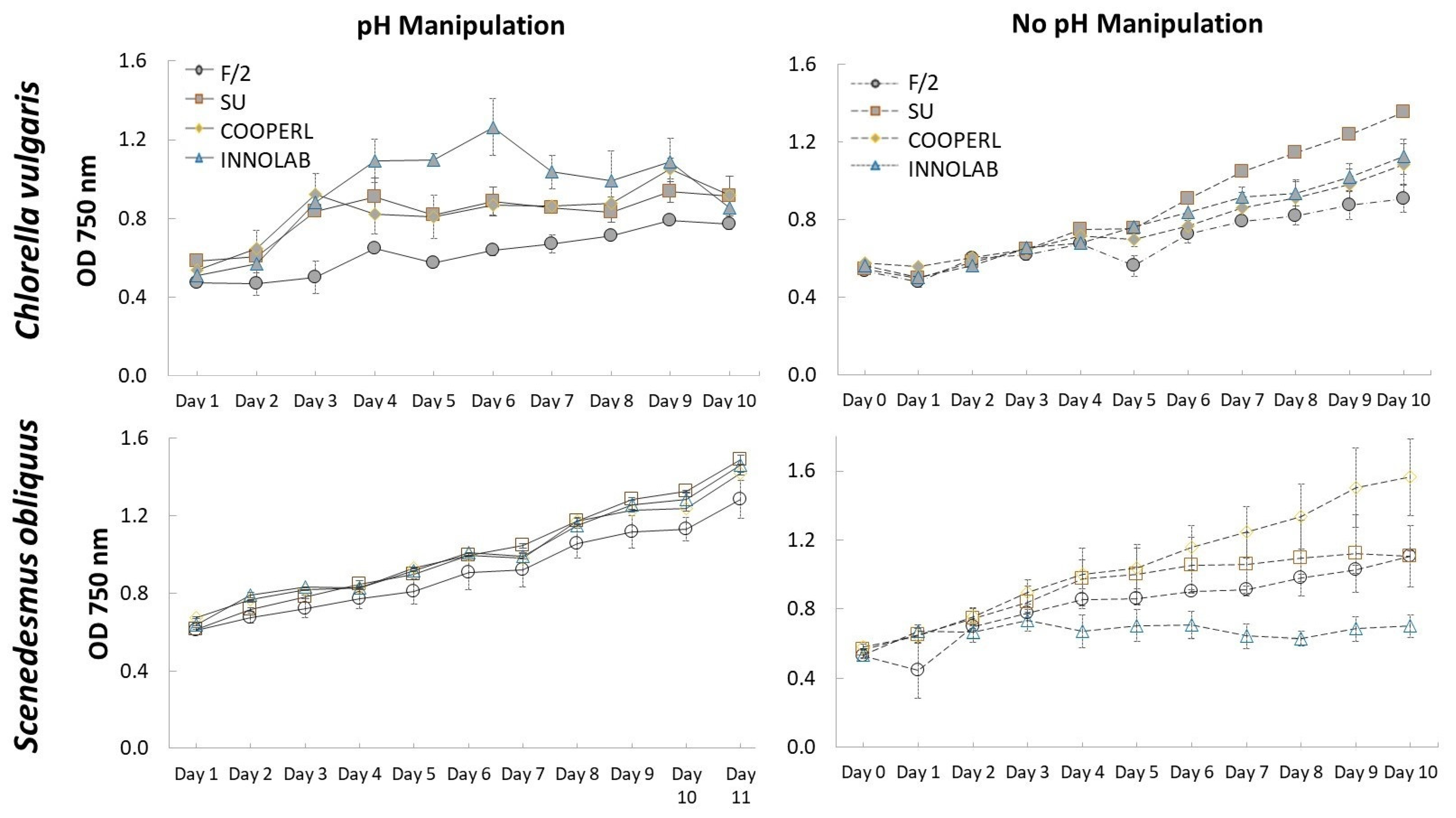
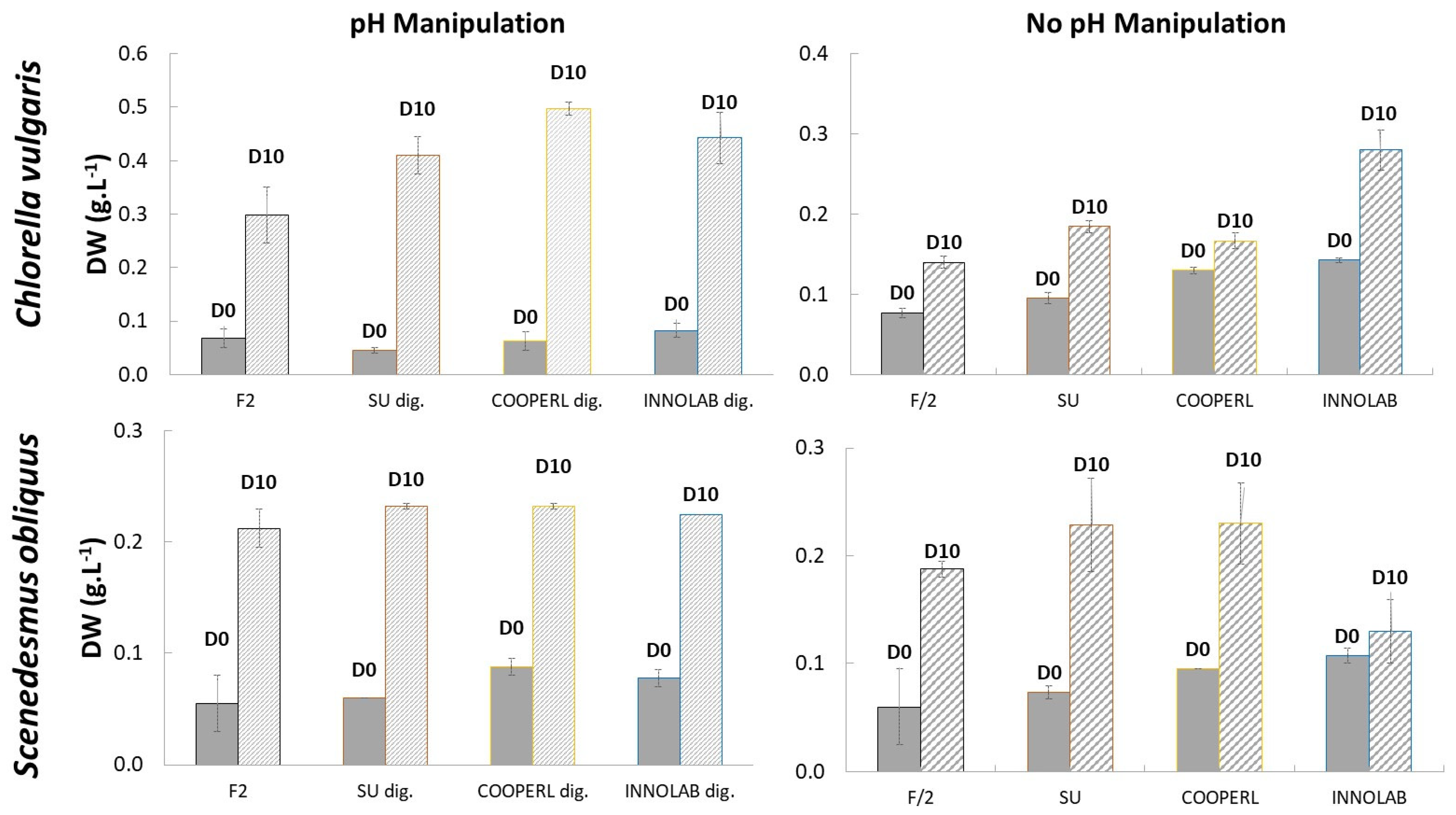
3.1. Growth Performances: Absorbance at 750 nm and Dry Weight
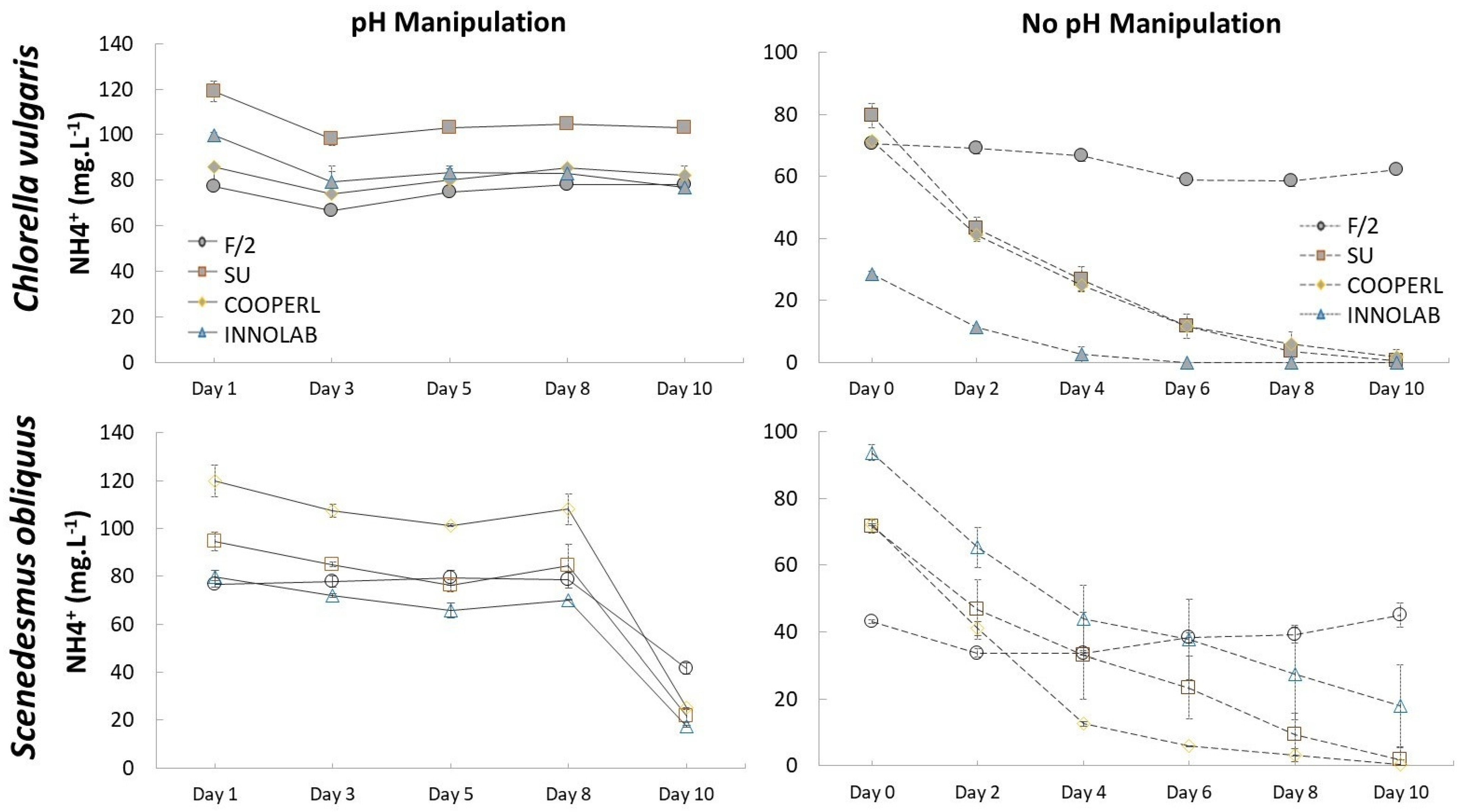
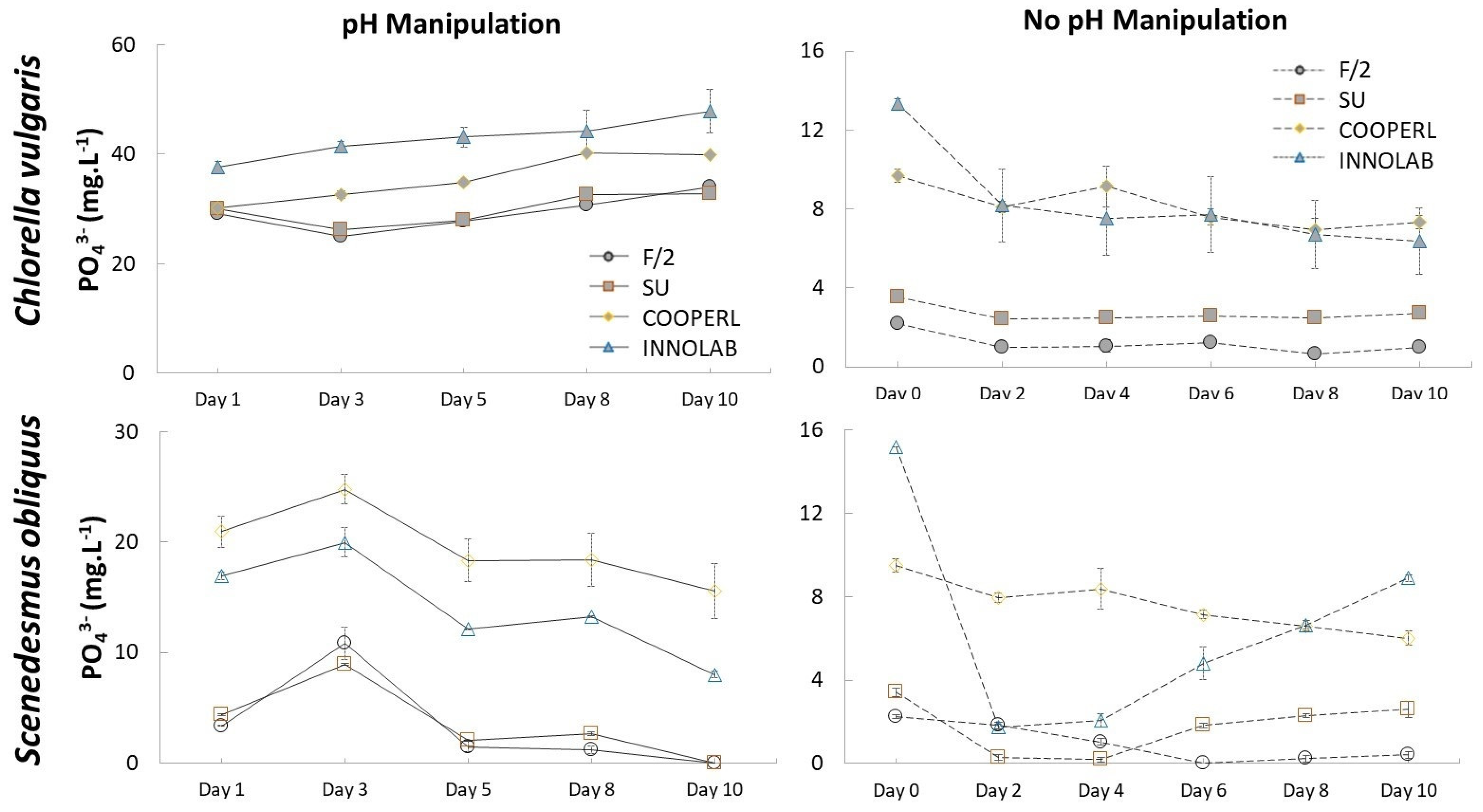
3.2. Nutrient Consumption: Ammonium and Phosphorus
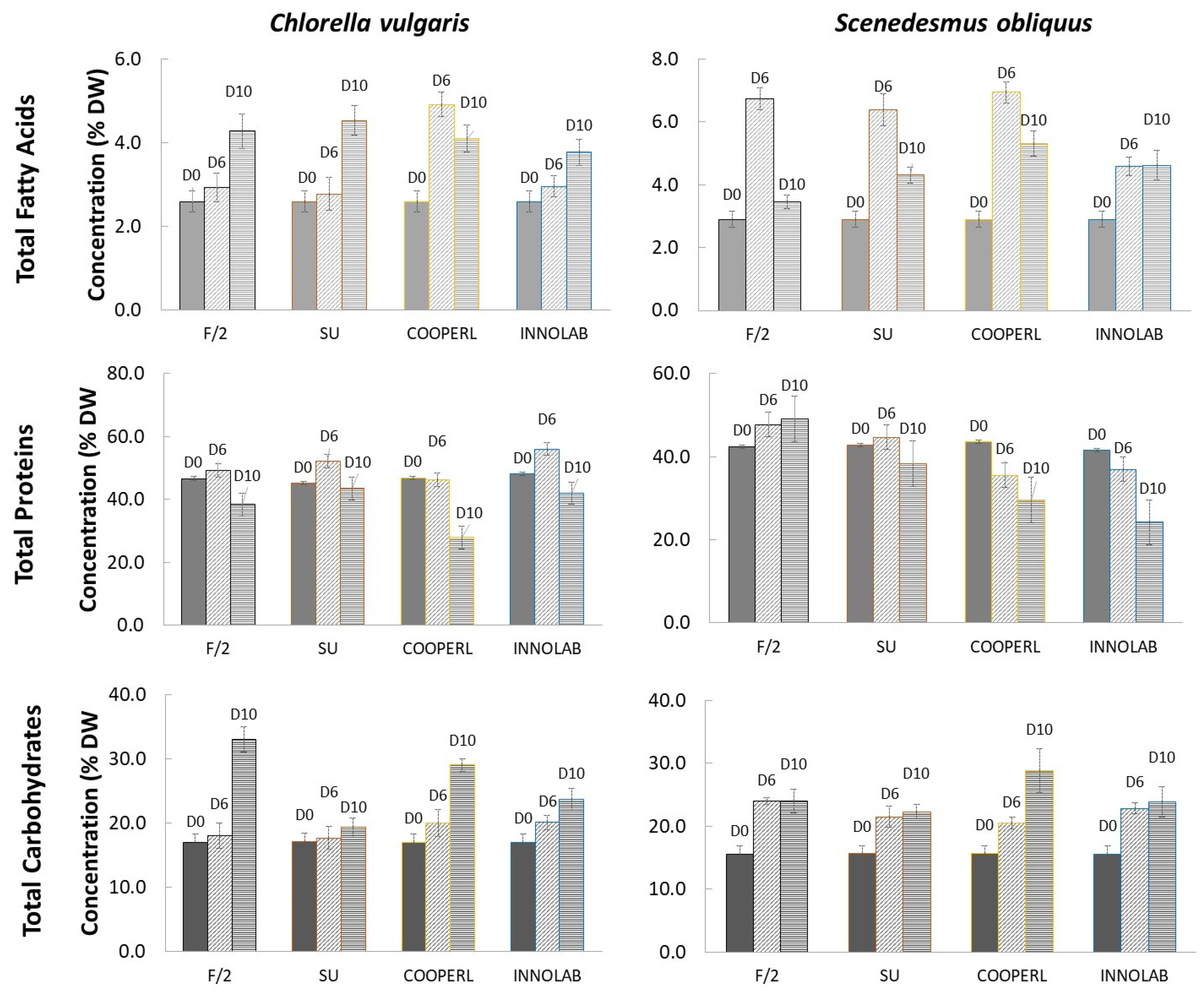
3.3. Composition Analysis: Fatty Acids, Proteins, and Carbohydrates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, W.; Drosg, B. Assessment of the State of the Art of Technologies for the Processing of Digestate Residue from Anaerobic Digesters. Water Sci. Technol. 2013, 67, 1984–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilayn, F.; Jimenez, J.; Martel, J.L.; Rouez, M.; Crest, M.; Patureau, D. First Fertilizing-Value Typology of Digestates: A Decision-Making Tool for Regulation. Waste Manag. 2019, 86, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.S.; Tan, W.G.; Halimatul Munawaroh, H.S.; Gupta, V.K.; Ho, S.H.; Show, P.L. Multifaceted Roles of Microalgae in the Application of Wastewater Biotreatment: A Review. Environ. Pollut. 2021, 269, 116236. [Google Scholar] [CrossRef] [PubMed]
- Wollmann, F.; Dietze, S.; Ackermann, J.U.; Bley, T.; Walther, T.; Steingroewer, J.; Krujatz, F. Microalgae Wastewater Treatment: Biological and Technological Approaches. Eng. Life Sci. 2019, 19, 860–871. [Google Scholar] [CrossRef] [Green Version]
- Silkina, A.; Ginnever, N.E.; Fernandes, F.; Fuentes-grünewald, C. Large-Scale Waste Bio-Remediation Using Microalgae Cultivation as a Platform. Energies 2019, 12, 2772. [Google Scholar] [CrossRef] [Green Version]
- Stiles, W.A.V.; Styles, D.; Chapman, S.P.; Esteves, S.; Bywater, A.; Melville, L.; Silkina, A.; Lupatsch, I.; Fuentes Grünewald, C.; Lovitt, R.; et al. Using Microalgae in the Circular Economy to Valorise Anaerobic Digestate: Challenges and Opportunities. Bioresour. Technol. 2018, 267, 732–742. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.C.; Cheng, Y.W.; Ishak, S.; Lam, M.K.; Lim, J.W.; Tan, I.S.; Show, P.L.; Lee, K.T. Anaerobic Digestate as a Low-Cost Nutrient Source for Sustainable Microalgae Cultivation: A Way Forward through Waste Valorization Approach. Sci. Total Environ. 2022, 803, 150070. [Google Scholar] [CrossRef]
- Silkina, A.; Zacharof, M.P.; Ginnever, N.E.; Gerardo, M.; Lovitt, R.W. Testing the Waste Based Biorefinery Concept: Pilot Scale Cultivation of Microalgal Species on Spent Anaerobic Digestate Fluids. Waste Biomass Valoriz. 2020, 11, 3883–3896. [Google Scholar] [CrossRef]
- Papadimitriou, E.K.; Barton, J.R.; Stentiford, E.I. Sources and Levels of Potentially Toxic Elements in the Biodegradable Fraction of Autoclaved Non-Segregated Household Waste and Its Compost/Digestate. Waste Manag. Res. 2008, 26, 419–430. [Google Scholar] [CrossRef]
- Tambone, F.; Orzi, V.; D’Imporzano, G.; Adani, F. Solid and Liquid Fractionation of Digestate: Mass Balance, Chemical Characterization, and Agronomic and Environmental Value. Bioresour. Technol. 2017, 243, 1251–1256. [Google Scholar] [CrossRef]
- Bauer, L.; Ranglová, K.; Masojídek, J.; Drosg, B.; Meixner, K. Digestate as Sustainable Nutrient Source for Microalgae—Challenges and Prospects. Appl. Sci. 2021, 11, 1056. [Google Scholar] [CrossRef]
- Fernandes, F.; Silkina, A.; Fuentes-Grünewald, C.; Wood, E.E.; Ndovela, V.L.S.; Oatley-Radcliffe, D.L.; Lovitt, R.W.; Llewellyn, C.A. Valorising Nutrient-Rich Digestate: Dilution, Settlement and Membrane Filtration Processing for Optimisation as a Waste-Based Media for Microalgal Cultivation. Waste Manag. 2020, 118, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Dębowski, M.; Szwaja, S.; Zieliński, M.; Kisielewska, M.; Stańczyk-Mazanek, E. The Influence of Anaerobic Digestion Effluents (ADEs) Used as the Nutrient Sources for Chlorella sp. Cultivation on Fermentative Biogas Production. Waste Biomass Valoriz. 2017, 8, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Barceló-Villalobos, M.; Gómez Serrano, C.; Sánchez Zurano, A.; Alameda García, L.; Esteve Maldonado, S.; Acién Fernández, J.F.G. Variations of culture parameters in a pilot-scale thin-layer reactor and their influence on the performance of Scenedesmus almeriensis culture. Bioresour. Technol. Rep. 2019, 6, 190–197. [Google Scholar] [CrossRef]
- Marcilhac, C.; Sialve, B.; Pourcher, A.M.; Ziebal, C.; Bernet, N.; Béline, F. Digestate Color and Light Intensity Affect Nutrient Removal and Competition Phenomena in a Microalgal-Bacterial Ecosystem. Water Res. 2014, 64, 278–287. [Google Scholar] [CrossRef]
- Lamolinara, B.; Pérez-Martínez, A.; Guardado-Yordi, E.; Guillén Fiallos, C.; Diéguez-Santana, K.; Ruiz-Mercado, G.J. Anaerobic Digestate Management, Environmental Impacts, and Techno-Economic Challenges. Waste Manag. 2022, 140, 14–30. [Google Scholar] [CrossRef]
- Häfner, F.; Hartung, J.; Möller, K. Digestate Composition Affecting N Fertiliser Value and C Mineralisation. Waste Biomass Valoriz. 2022. [Google Scholar] [CrossRef]
- Salbitani, G.; Carfagna, S. Ammonium Utilization in Microalgae: A Sustainable Method for Wastewater Treatment. Sustainability 2021, 13, 956. [Google Scholar] [CrossRef]
- Mandal, S.; Shurin, J.B.; Efroymson, R.A.; Mathews, T.J. Functional Divergence in Nitrogen Uptake Rates Explains Diversity–Productivity Relationship in Microalgal Communities. Ecosphere 2018, 9, e02228. [Google Scholar] [CrossRef]
- Shen, Y.; Qiu, S.; Chen, Z.; Zhang, Y.; Trent, J.; Ge, S. Free Ammonia Is the Primary Stress Factor Rather than Total Ammonium to Chlorella sorokiniana in Simulated Sludge Fermentation Liquor. Chem. Eng. J. 2020, 397, 125490. [Google Scholar] [CrossRef]
- Chuka-ogwude, D.; Ogbonna, J.; Borowitzka, M.A.; Moheimani, N.R. Screening, Acclimation and Ammonia Tolerance of Microalgae Grown in Food Waste Digestate. J. Appl. Phycol. 2020, 32, 3775–3785. [Google Scholar] [CrossRef]
- Chanda, M.J.; Merghoub, N.; El Arroussi, H. Microalgae Polysaccharides: The New Sustainable Bioactive Products for the Development of Plant Bio-Stimulants? World J. Microbiol. Biotechnol. 2019, 35, 177. [Google Scholar] [CrossRef] [PubMed]
- Kapoore, R.V.; Wood, E.E.; Llewellyn, C.A. Algae Biostimulants: A Critical Look at Microalgal Biostimulants for Sustainable Agricultural Practices. Biotechnol. Adv. 2021, 49, 107754. [Google Scholar] [CrossRef]
- Chuka-ogwude, D.; Ogbonna, J.; Moheimani, N.R. A Review on Microalgal Culture to Treat Anaerobic Digestate Food Waste Effluent. Algal Res. 2020, 47, 101841. [Google Scholar] [CrossRef]
- Zieliński, M.; Dębowski, M.; Kazimierowicz, J. Outflow from a Biogas Plant as a Medium for Microalgae Biomass Cultivation—Pilot Scale Study and Technical Concept of a Large-Scale Installation. Energies 2022, 15, 2912. [Google Scholar] [CrossRef]
- Judd, S.; van den Broeke, L.J.P.; Shurair, M.; Kuti, Y.; Znad, H. Algal Remediation of CO2and Nutrient Discharges: A Review. Water Res. 2015, 87, 356–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chioccioli, M.; Hankamer, B.; Ross, I.L. Flow Cytometry Pulse Width Data Enables Rapid and Sensitive Estimation of Biomass Dry Weight in the Microalgae Chlamydomonas reinhardtii and Chlorella vulgaris. PLoS ONE 2014, 9, e97269. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Vaidyanathan, S. Simultaneous Assay of Pigments, Carbohydrates, Proteins and Lipids in Microalgae. Anal. Chim. Acta 2013, 776, 31–40. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Huete-Ortega, M.; Day, J.G.; Okurowska, K.; Slocombe, S.P.; Stanley, M.S.; Vaidyanathan, S. Effects of Cryopreservation on Viability and Functional Stability of an Industrially Relevant Alga. Sci. Rep. 2019, 9, 2093. [Google Scholar] [CrossRef]
- Hamed, S.M.; Kapoore, R.V.; Raut, M.P.; Vaidyanathan, S.; Wright, P.C. Influence of Nutrient Status on the Biohydrogen and Lipid Productivity in Parachlorella Kessleri: A Biorefinery Approach. Appl. Microbiol. Biotechnol. 2020, 104, 10293–10305. [Google Scholar] [CrossRef]
- Kapoore, R.V. Mass Spectrometry Based Hyphenated Techniques for Microalgal and Mammalian Metabolomics. Ph.D. Thesis, University of Sheffield, Sheffield, UK, August 2014. [Google Scholar]
- Roccuzzo, S.; Couto, N.; Karunakaran, E.; Kapoore, R.V.; Butler, T.O.; Mukherjee, J.; Hansson, E.M.; Beckerman, A.P.; Pandhal, J. Metabolic Insights Into Infochemicals Induced Colony Formation and Flocculation in Scenedesmus subspicatus Unraveled by Quantitative Proteomics. Front. Microbiol. 2020, 11, 792. [Google Scholar] [CrossRef] [PubMed]
- Chuka-ogwude, D.; Ogbonna, J.C.; Moheimani, N.R. Adjustments of the Photosynthetic Unit and Compensation Mechanisms of Tolerance to High Ammonia Concentration in Chlorella sp. Grown in Food Waste Digestate. Algal Res. 2020, 52, 102106. [Google Scholar] [CrossRef]
- Su, Y. Revisiting Carbon, Nitrogen, and Phosphorus Metabolisms in Microalgae for Wastewater Treatment. Sci. Total Environ. 2021, 762, 144590. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Shilton, A. Luxury Uptake of Phosphorus by Microalgae in Waste Stabilisation Ponds: Current Understanding and Future Direction. Rev. Environ. Sci. Biotechnol. 2014, 13, 321–328. [Google Scholar] [CrossRef]
- Gerardo, M.L.; Aljohani, N.H.M.; Oatley-radcliffe, D.L.; Lovitt, R.W. Moving towards Sustainable Resources: Recovery and Fractionation of Nutrients from Dairy Manure Digestate Using Membranes. Water Res. 2015, 80, 80–89. [Google Scholar] [CrossRef]
- Su, J.F.; Ma, M.; Wei, L.; Ma, F.; Lu, J.S.; Shao, S.C. Algicidal and Denitrification Characterization of Acinetobacter sp. J25 against Microcystis aeruginosa and Microbial Community in Eutrophic Landscape Water. Mar. Pollut. Bull. 2016, 107, 233–239. [Google Scholar] [CrossRef]
- Mukherjee, C.; Chowdhury, R.; Ray, K. Phosphorus Recycling from an Unexplored Source by Polyphosphate Accumulating Microalgae and Cyanobacteria—A Step to Phosphorus Security in Agriculture. Front. Microbiol. 2015, 6, 1421. [Google Scholar] [CrossRef] [Green Version]
- Sforza, E.; Calvaruso, C.; La Rocca, N.; Bertucco, A. Luxury Uptake of Phosphorus in Nannochloropsis salina: Effect of P Concentration and Light on P Uptake in Batch and Continuous Cultures. Biochem. Eng. J. 2018, 134, 69–79. [Google Scholar] [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Selyakh, I.; Semenova, L.; Ismagulova, T.; Lukyanov, A.; Mamedov, I.; Vinogradova, E.; Karpova, O.; Konyukhov, I.; et al. Phosphorus Starvation and Luxury Uptake in Green Microalgae Revisited. Algal Res. 2019, 43, 101651. [Google Scholar] [CrossRef]
- John, E.H.; Flynn, K.J. Modelling Phosphate Transport and Assimilation in Microalgae; How Much Complexity Is Warranted? Ecol. Model. 2000, 125, 145–157. [Google Scholar] [CrossRef]
- Eixler, S.; Karsten, U.; Selig, U. Phosphorus Storage in Chlorella vulgaris (Trebouxiophyceae, Chlorophyta) Cells and Its Dependence on Phosphate Supply. Phycologia 2006, 45, 53–60. [Google Scholar] [CrossRef]
- Nishikawa, K.; Yamakoshi, Y.; Uemura, I.; Tominaga, N. Ultrastructural Changes in Chlamydomonas acidophila (Chlorophyta) Induced by Heavy Metals and Polyphosphate Metabolism. FEMS Microbiol. Ecol. 2003, 44, 253–259. [Google Scholar] [CrossRef]
- Powell, N.; Shilton, A.; Pratt, S.; Chisti, Y. Luxury Uptake of Phosphorus by Microalgae in Full-Scale Waste Stabilisation Ponds. Water Sci. Technol. 2011, 63, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of Carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Bouras, S.; Antoniadis, D.; Kountrias, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of PH on Schizochytrium limacinum Production Grown Using Crude Glycerol and Biogas Digestate Effluent. Agronomy 2022, 12, 364. [Google Scholar] [CrossRef]
- Khanh, N.; Kitaya, Y.; Xiao, L.; Endo, R.; Shibuya, T. Selection of Microalgae Suitable for Culturing with Digestate from Methane Fermentation. Environ. Technol. 2013, 34, 2039–2045. [Google Scholar] [CrossRef]
- Al-Mallahi, J.; Ishii, K. Attempts to Alleviate Inhibitory Factors of Anaerobic Digestate for Enhanced Microalgae Cultivation and Nutrients Removal: A Review. J. Environ. Manag. 2022, 304, 114266. [Google Scholar] [CrossRef]
- Jiang, R.; Qin, L.; Feng, S.; Huang, D.; Wang, Z.; Zhu, S. The Joint Effect of Ammonium and pH on the Growth of Chlorella vulgaris and Ammonium Removal in Artificial Liquid Digestate. Bioresour. Technol. 2021, 325, 124690. [Google Scholar] [CrossRef]
- Park, J.; Jin, H.F.; Lim, B.R.; Park, K.Y.; Lee, K. Ammonia Removal from Anaerobic Digestion Effluent of Livestock Waste Using Green Alga Scenedesmus sp. Bioresour. Technol. 2010, 101, 8649–8657. [Google Scholar] [CrossRef]
- Wang, Q.; Hyman, M.; Higgins, B.T. Factors Impacting the Effectiveness of Biological Pretreatment for the Alleviation of Algal Growth Inhibition on Anaerobic Digestate. Algal Res. 2021, 53, 102129. [Google Scholar] [CrossRef]
- Zuliani, L.; Frison, N.; Jelic, A.; Fatone, F.; Bolzonella, D.; Ballottari, M. Microalgae Cultivation on Anaerobic Digestate of Municipal Wastewater, Sewage Sludge and Agro-Waste. Int. J. Mol. Sci. 2016, 17, 1692. [Google Scholar] [CrossRef] [PubMed]
- Cheunbarn, S.; Peerapornpisal, Y. Cultivation of Spirulina platensis Using Anaerobically Swine Wastewater Treatment Effluent. Int. J. Agric. Biol. 2010, 12, 586–590. [Google Scholar]
- Uggetti, E.; Sialve, B.; Latrille, E.; Steyer, J.-P. Anaerobic Digestate as Substrate for Microalgae Culture: The Role of Ammonium Concentration on the Microalgae Productivity. Bioresour. Technol. 2014, 152, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.S.G.; van den Berg, E.; Stieger, M. The Influence of Product Preparation, Familiarity and Individual Traits on the Consumer Acceptance of Insects as Food. Food Qual. Prefer. 2016, 52, 222–231. [Google Scholar] [CrossRef]
- Wang, L.; Li, Y.; Chen, P.; Min, M.; Chen, Y.; Zhu, J.; Ruan, R.R. Anaerobic Digested Dairy Manure as a Nutrient Supplement for Cultivation of Oil-Rich Green Microalgae Chlorella sp. Bioresour. Technol. 2010, 101, 2623–2628. [Google Scholar] [CrossRef] [PubMed]
- Koutra, E.; Grammatikopoulos, G.; Kornaros, M. Selection of Microalgae Intended for Valorization of Digestate from Agro-Waste Mixtures. Waste Manag. 2018, 73, 123–129. [Google Scholar] [CrossRef]
- Olguín, E.J.; Dorantes, E.; Castillo, O.S.; Hernández-Landa, V.J. Anaerobic Digestates from Vinasse Promote Growth and Lipid Enrichment in Neochloris oleoabundans Cultures. J. Appl. Phycol. 2015, 27, 1813–1822. [Google Scholar] [CrossRef]
- Cai, T.; Ge, X.; Park, S.Y.; Li, Y. Comparison of Synechocystis sp. PCC6803 and Nannochloropsis salina for Lipid Production Using Artificial Seawater and Nutrients from Anaerobic Digestion Effluent. Bioresour. Technol. 2013, 144, 255–260. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, S.; Mo, N.; Wang, Z.; Zeng, E.Y. Screening of Freshwater Oleaginous Microalgae from South China and Its Cultivation Characteristics in Energy Grass Digestate. J. Clean. Prod. 2020, 276, 124193. [Google Scholar] [CrossRef]
- Michelon, W.; Da Silva, M.L.B.; Mezzari, M.P.; Pirolli, M.; Prandini, J.M.; Soares, H.M. Effects of Nitrogen and Phosphorus on Biochemical Composition of Microalgae Polyculture Harvested from Phycoremediation of Piggery Wastewater Digestate. Appl. Biochem. Biotechnol. 2016, 178, 1407–1419. [Google Scholar] [CrossRef]
- Kumar, A.; Bera, S. Revisiting Nitrogen Utilization in Algae: A Review on the Process of Regulation and Assimilation. Bioresour. Technol. Rep. 2020, 12, 100584. [Google Scholar] [CrossRef]
- Massa, M.; Buono, S.; Langellotti, A.L.; Castaldo, L.; Martello, A.; Paduano, A.; Sacchi, R.; Fogliano, V. Evaluation of Anaerobic Digestates from Different Feedstocks as Growth Media for Tetradesmus obliquus, Botryococcus braunii, Phaeodactylum tricornutum and Arthrospira maxima. New Biotechnol. 2017, 36, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Nwoba, E.G.; Ayre, J.M.; Moheimani, N.R.; Ubi, B.E.; Ogbonna, J.C. Growth Comparison of Microalgae in Tubular Photobioreactor and Open Pond for Treating Anaerobic Digestion Piggery Effluent. Algal Res. 2016, 17, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Depraetere, O.; Deschoenmaeker, F.; Badri, H.; Monsieurs, P.; Foubert, I.; Leys, N.; Wattiez, R.; Muylaert, K. Trade-off between Growth and Carbohydrate Accumulation in Nutrient-Limited Arthrospira sp. PCC 8005 Studied by Integrating Transcriptomic and Proteomic Approaches. PLoS ONE 2015, 10, e0132461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledda, C.; Schievano, A.; Scaglia, B.; Rossoni, M.; Acién Fernández, F.G.; Adani, F. Integration of Microalgae Production with Anaerobic Digestion of Dairy Cattle Manure: An Overall Mass and Energy Balance of the Process. J. Clean. Prod. 2016, 112, 103–112. [Google Scholar] [CrossRef]
- Huo, Y.X.; Cho, K.M.; Rivera, J.G.L.; Monte, E.; Shen, C.R.; Yan, Y.; Liao, J.C. Conversion of Proteins into Biofuels by Engineering Nitrogen Flux. Nat. Biotechnol. 2011, 29, 346–351. [Google Scholar] [CrossRef]
- BenMoussa-Dahmen, I.; Chtourou, H.; Rezgui, F.; Sayadi, S.; Dhouib, A. Salinity Stress Increases Lipid, Secondary Metabolites and Enzyme Activity in Amphora Subtropica and Dunaliella sp. for Biodiesel Production. Bioresour. Technol. 2016, 218, 816–825. [Google Scholar] [CrossRef]
- Brányiková, I.; Maršálková, B.; Doucha, J.; Brányik, T.; Bišová, K.; Zachleder, V.; Vítová, M. Microalgae-Novel Highly Efficient Starch Producers. Biotechnol. Bioeng. 2011, 108, 766–776. [Google Scholar] [CrossRef]
- Ji, C.F.; Yu, X.J.; Chen, Z.A.; Xue, S.; Legrand, J.; Zhang, W. Effects of Nutrient Deprivation on Biochemical Compositions and Photo-Hydrogen Production of Tetraselmis Subcordiformis. Int. J. Hydrogen Energy 2011, 36, 5817–5821. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NH4+ | PO43− | N:P | Dilution Factor Required | |
|---|---|---|---|---|
| SU | 4815 mg L−1 | 202 mg L−1 | 23.8 | 48 |
| COOPERL | 3102 mg L−1 | 440 mg L−1 | 7.1 | 31 |
| INNOLAB | 5295 mg L−1 | 395 mg L−1 | 13.4 | 53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, F.; Silkina, A.; Gayo-Peláez, J.I.; Kapoore, R.V.; de la Broise, D.; Llewellyn, C.A. Microalgae Cultivation on Nutrient Rich Digestate: The Importance of Strain and Digestate Tailoring under PH Control. Appl. Sci. 2022, 12, 5429. https://doi.org/10.3390/app12115429
Fernandes F, Silkina A, Gayo-Peláez JI, Kapoore RV, de la Broise D, Llewellyn CA. Microalgae Cultivation on Nutrient Rich Digestate: The Importance of Strain and Digestate Tailoring under PH Control. Applied Sciences. 2022; 12(11):5429. https://doi.org/10.3390/app12115429
Chicago/Turabian StyleFernandes, Fleuriane, Alla Silkina, José Ignacio Gayo-Peláez, Rahul Vijay Kapoore, Denis de la Broise, and Carole A. Llewellyn. 2022. "Microalgae Cultivation on Nutrient Rich Digestate: The Importance of Strain and Digestate Tailoring under PH Control" Applied Sciences 12, no. 11: 5429. https://doi.org/10.3390/app12115429