FOXL2 is a Progesterone Target Gene in the Endometrium of Ruminants

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. FOXL2 is Expressed in the Ovine Endometrium
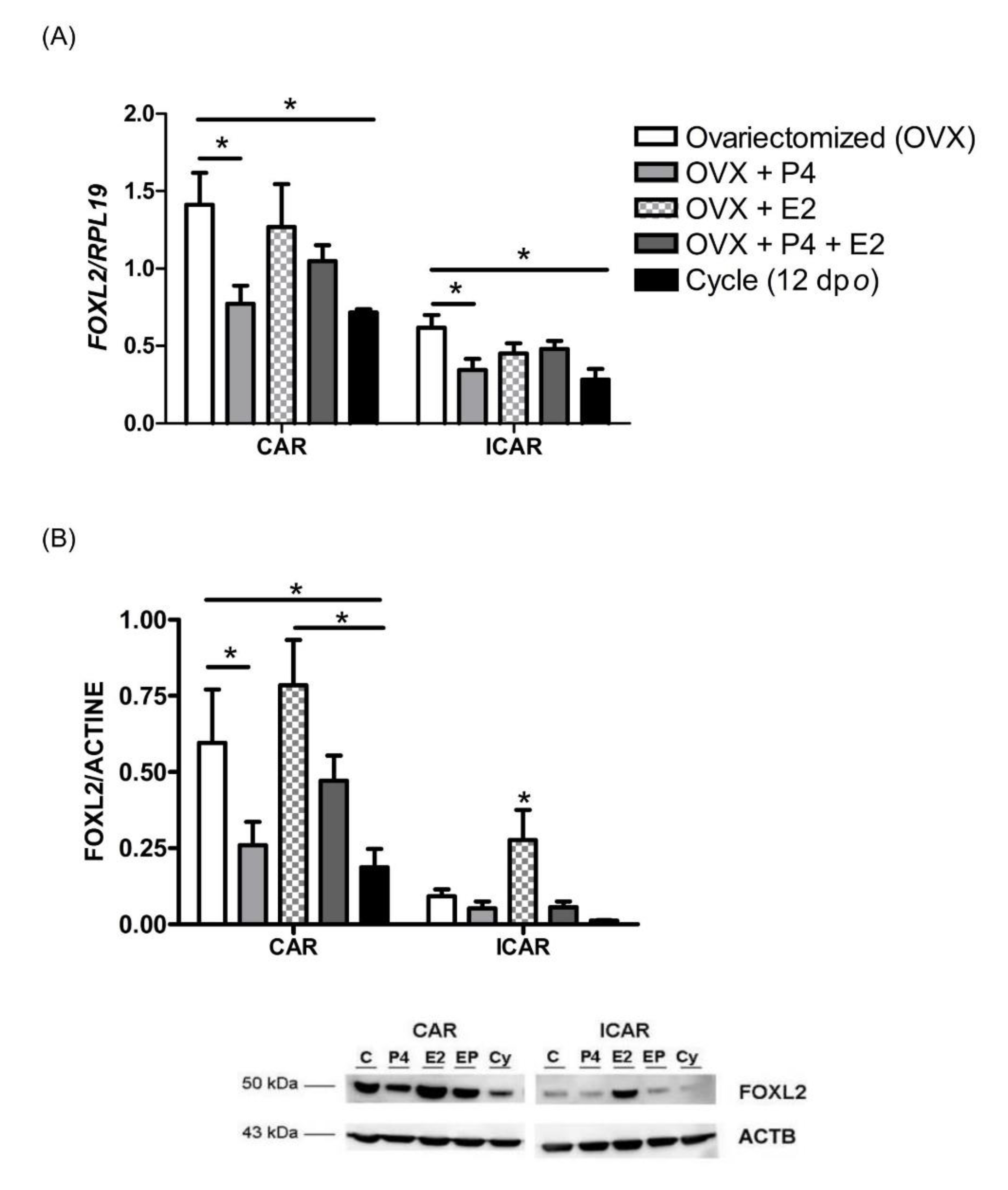
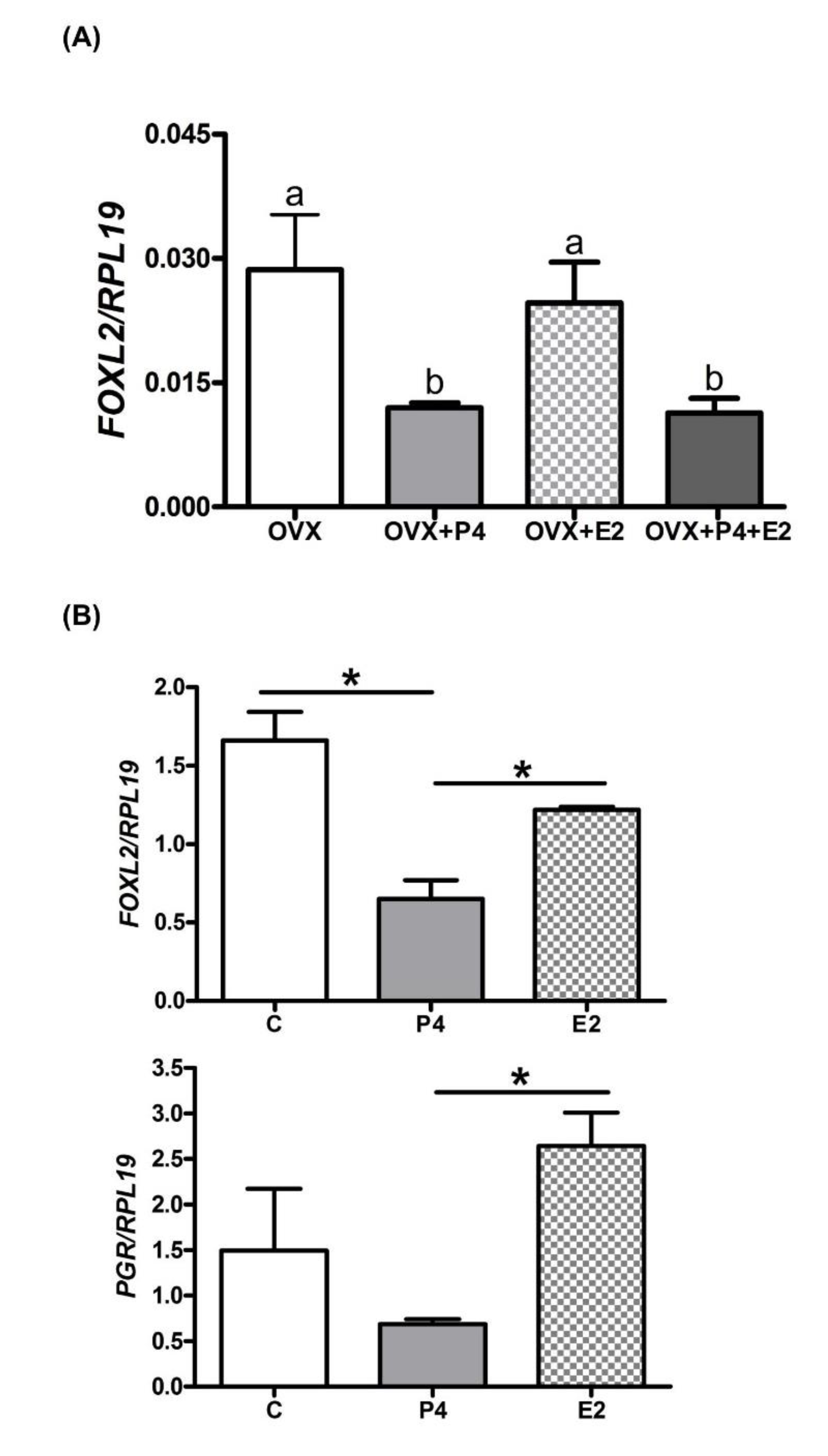
2.2. Endometrial FOXL2 Gene Expression Varies with Blood P4 Concentrations in Sheep
2.3. P4 Supplementation Reduces Endometrial FOXL2 Gene Expression in Cattle
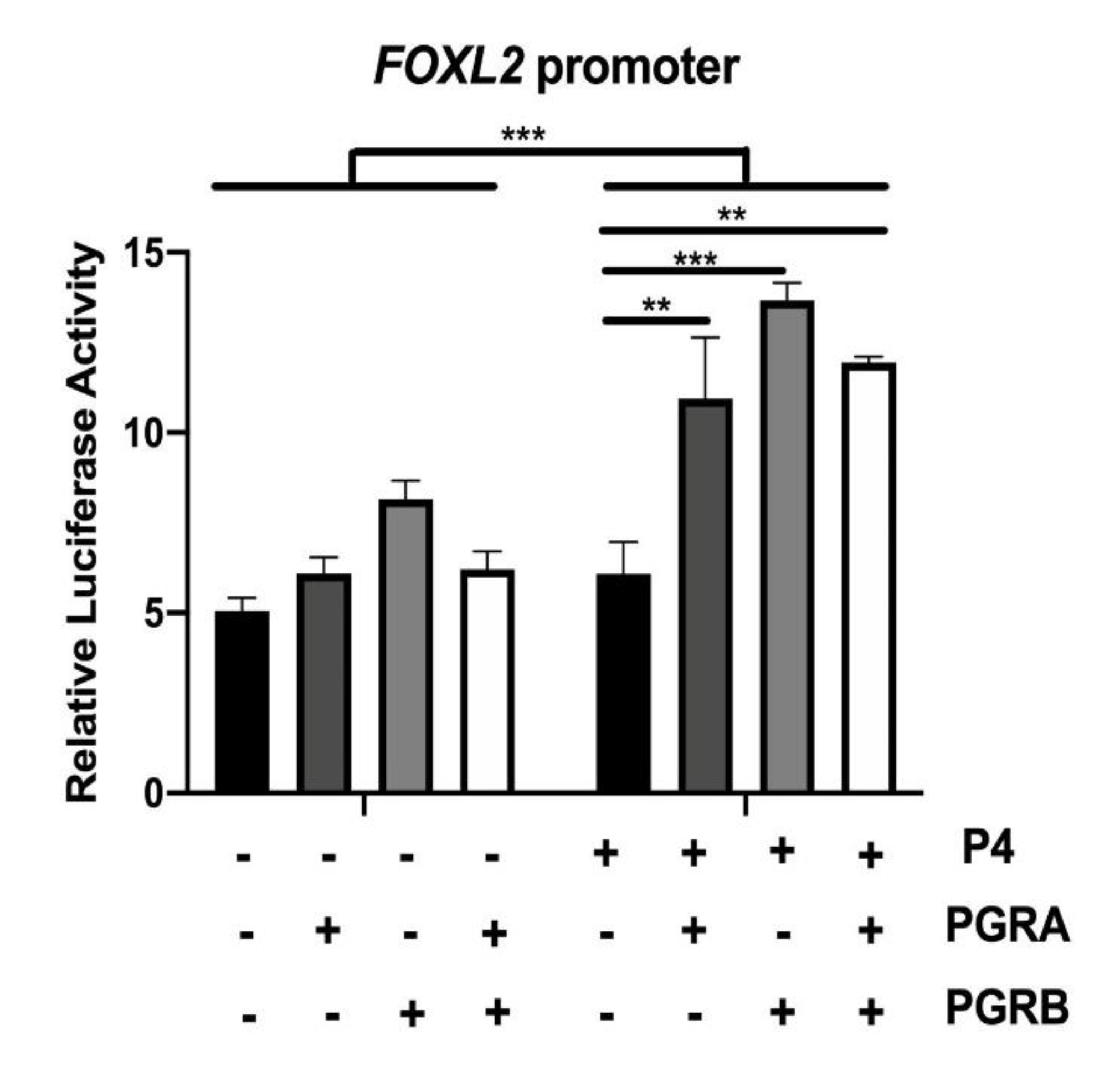
2.4. P4 Directly Regulates the Activity of FOXL2 Promoter through Its Nuclear Receptor
3. Discussion
4. Materials and Methods
4.1. Animal Experiments and Cell Cultures
4.2. In Vivo Supplementation of Steroids in Ewes
4.3. In Vivo Supplementation of Steroids in Cows
4.4. Incubation of Bovine Endometrial Explants with Steroids
4.5. Cell Culture Conditions and Transfections Assays in COS Cells
4.6. Tissue and Cell Collection and RNA Extraction
4.7. Real-Time RT-PCR
4.8. Western Blot Analysis
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bazer, F.W.; Wu, G.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C.; Bayless, K. Novel pathways for implantation and establishment and maintenance of pregnancy in mammals. Mol. Hum. Reprod. 2010, 16, 135–152. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; DeMayo, F.J. Animal models of implantation. Reproduction 2004, 128, 679–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandra, O.; Charpigny, G.; Galio, L.; Hue, I. Preattachment Embryos of Domestic Animals: Insights into Development and Paracrine Secretions. Annu. Rev. Anim. Biosci. 2017, 5, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Hansen, T.R. Implantation and Establishment of Pregnancy in Ruminants. Adv. Anat. Embryol. Cell Biol. 2015, 216, 105–135. [Google Scholar] [PubMed]
- Franczyk, M.; Lopucki, M.; Stachowicz, N.; Morawska, D.; Kankofer, M. Extracellular matrix proteins in healthy and retained placentas, comparing hemochorial and synepitheliochorial placentas. Placenta 2017, 50, 19–24. [Google Scholar] [CrossRef]
- Hafez, S. Comparative Placental Anatomy: Divergent Structures Serving a Common Purpose. Prog. Mol. Biol. Transl. Sci. 2017, 145, 1–28. [Google Scholar] [PubMed]
- Sandra, O.; Mansouri-Attia, N.; Lea, R.G. Novel aspects of endometrial function: A biological sensor of embryo quality and driver of pregnancy success. Reprod. Fertil. Dev. 2011, 24, 68–79. [Google Scholar] [CrossRef]
- Amoako, A.A.; Nafee, T.M.; Ola, B. Epigenetic Influences During the Periconception Period and Assisted Reproduction. Adv. Exp. Med. Biol. 2017, 1014, 15–39. [Google Scholar]
- Diskin, M.G.; Morris, D.G. Embryonic and early foetal losses in cattle and other ruminants. Reprod. Domest. Anim. 2008, 43 (Suppl. 2), 260–267. [Google Scholar] [CrossRef]
- Wiltbank, M.C.; Baez, G.M.; Garcia-Guerra, A.; Toledo, M.Z.; Monteiro, P.L.; Melo, L.F.; Ochoa, J.C.; Santos, J.E.; Sartori, R. Pivotal periods for pregnancy loss during the first trimester of gestation in lactating dairy cows. Theriogenology 2016, 86, 239–253. [Google Scholar] [CrossRef]
- Al-Gubory, K.H.; Bolifraud, P.; Garrel, C. Regulation of Key Antioxidant Enzymatic Systems in the Sheep Endometrium by Ovarian Steroids. Endocrinology 2008, 149, 4428–4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devroey, P.; Pados, G. Preparation of endometrium for egg donation. Hum. Reprod. Update 1998, 4, 856–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, B.G.; Moore, N.W. Effects of progesterone and oestradiol on endometrial metabolism and embryo survival in the ovariectomized ewe. Theriogenology 1976, 6, 636. [Google Scholar] [CrossRef]
- Ozturk, S.; Demir, R. Particular functions of estrogen and progesterone in establishment of uterine receptivity and embryo implantation. Histol. Histopathol. 2010, 25, 1215–1228. [Google Scholar]
- Patel, B.; Elguero, S.; Thakore, S.; Dahoud, W.; Bedaiwy, M.; Mesiano, S. Role of nuclear progesterone receptor isoforms in uterine pathophysiology. Hum. Reprod. Update 2015, 21, 155–173. [Google Scholar] [CrossRef] [Green Version]
- Qu, T.; Zhang, S.M.; Yu, L.L.; Zhang, S.; Yuan, D.Z.; Xu, Q.; Zhang, J.H.; He, Y.P.; Yue, L.M. Relocalisation and activation of integrins induced rapidly by oestrogen via G-protein-coupled receptor 30 in mouse blastocysts. Reprod. Fertil. Dev. 2015. [Google Scholar] [CrossRef]
- Szekeres-Bartho, J.; Sucurovic, S.; Mulac-Jericevic, B. The Role of Extracellular Vesicles and PIBF in Embryo-Maternal Immune-Interactions. Front. Immunol. 2018, 9, 2890. [Google Scholar] [CrossRef]
- Forde, N.; Beltman, M.E.; Lonergan, P.; Diskin, M.; Roche, J.F.; Crowe, M.A. Oestrous cycles in Bos taurus cattle. Anim. Reprod. Sci. 2011, 124, 163–169. [Google Scholar] [CrossRef]
- Spencer, T.E.; Forde, N.; Lonergan, P. Insights into conceptus elongation and establishment of pregnancy in ruminants. Reprod. Fertil. Dev. 2016, 29, 84–100. [Google Scholar] [CrossRef]
- De Ruijter-Villani, M.; Deelen, C.; Stout, T.A. Expression of leukaemia inhibitory factor at the conceptus?maternal interface during preimplantation development and in the endometrium during the oestrous cycle in the mare. Reprod. Fertil. Dev. 2015. [Google Scholar] [CrossRef]
- Hawkins, S.M.; Matzuk, M.M. The menstrual cycle: Basic biology. Ann. N. Y. Acad. Sci. 2008, 1135, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Mihm, M.; Gangooly, S.; Muttukrishna, S. The normal menstrual cycle in women. Anim. Reprod. Sci. 2011, 124, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P. Influence of progesterone on oocyte quality and embryo development in cows. Theriogenology 2011, 76, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Spencer, T.E.; Johnson, G.A.; Burghardt, R.C.; Wu, G. Comparative aspects of implantation. Reproduction 2009, 138, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Forde, N.; Lonergan, P. Transcriptomic analysis of the bovine endometrium: What is required to establish uterine receptivity to implantation in cattle? J. Reprod. Dev. 2012, 58, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.M.; Randi, F.; Passaro, C.; Mathew, D.J.; Butler, S.T.; Lonergan, P. Effect of human chorionic gonadotrophin administration 2 days after insemination on progesterone concentration and pregnancy per artificial insemination in lactating dairy cows. J. Dairy Sci. 2018, 101, 6556–6567. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C.; Palmarini, M. Pregnancy recognition and conceptus implantation in domestic ruminants: Roles of progesterone, interferons and endogenous retroviruses. Reprod. Fertil. Dev. 2007, 19, 65–78. [Google Scholar] [CrossRef]
- Martal, J.; Lacroix, M.C.; Loudes, C.; Saunier, M.; Wintenberger-Torres, S. Trophoblastin, an antiluteolytic protein present in early pregnancy in sheep. J. Reprod. Fertil. 1979, 56, 63–73. [Google Scholar] [CrossRef]
- Roberts, R.M.; Chen, Y.; Ezashi, T.; Walker, A.M. Interferons and the maternal-conceptus dialog in mammals. Semin. Cell Dev. Biol. 2008, 19, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Sandra, O.; Wolf, E. Genes involved in conceptus-endometrial interactions in ruminants: Insights from reductionism and thoughts on holistic approaches. Reproduction 2008, 135, 165–179. [Google Scholar] [CrossRef]
- Hansen, T.R.; Sinedino, L.D.P.; Spencer, T.E. Paracrine and endocrine actions of interferon tau (IFNT). Reproduction 2017, 154, F45–F59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imakawa, K.; Bai, R.; Kusama, K. Integration of molecules to construct the processes of conceptus implantation to the maternal endometrium. J. Anim. Sci. 2018, 96, 3009–3021. [Google Scholar] [CrossRef]
- Lonergan, P.; Forde, N. The Role of Progesterone in Maternal Recognition of Pregnancy in Domestic Ruminants. Adv. Anat. Embryol. Cell Biol. 2015, 216, 87–104. [Google Scholar]
- Wetendorf, M.; DeMayo, F.J. Progesterone receptor signaling in the initiation of pregnancy and preservation of a healthy uterus. Int. J. Dev. Biol. 2014, 58, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, R.M.; Kim, T.H.; Shin, J.H.; Jeong, J.W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, M.; de La Fuente, J.; Fair, T.; Al Naib, A.; Gutierrez-Adan, A.; Roche, J.F.; Rizos, D.; Lonergan, P. Progesterone and conceptus elongation in cattle: A direct effect on the embryo or an indirect effect via the endometrium? Reproduction 2009, 138, 507–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumu, L.A.; Forde, N.; Fahey, A.G.; Fitzpatrick, E.; Roche, J.F.; Crowe, M.A.; Lonergan, P. The effect of elevated progesterone and pregnancy status on mRNA expression and localisation of progesterone and oestrogen receptors in the bovine uterus. Reproduction 2010, 140, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wu, S.P.; DeMayo, F.J. Hormone dependent uterine epithelial-stromal communication for pregnancy support. Placenta 2017, 60 (Suppl. 1), S20–S26. [Google Scholar] [CrossRef]
- Wetendorf, M.; Wu, S.P.; Wang, X.; Creighton, C.J.; Wang, T.; Lanz, R.B.; Blok, L.; Tsai, S.Y.; Tsai, M.J.; Lydon, J.P.; et al. Decreased epithelial progesterone receptor A at the window of receptivity is required for preparation of the endometrium for embryo attachment. Biol. Reprod. 2017, 96, 313–326. [Google Scholar] [CrossRef]
- Vasquez, Y.M.; Mazur, E.C.; Li, X.; Kommagani, R.; Jiang, L.; Chen, R.; Lanz, R.B.; Kovanci, E.; Gibbons, W.E.; DeMayo, F.J. FOXO1 is required for binding of PR on IRF4, novel transcriptional regulator of endometrial stromal decidualization. Mol. Endocrinol. 2015, 29, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Bauersachs, S.; Mitko, K.; Ulbrich, S.E.; Blum, H.; Wolf, E. Transcriptome studies of bovine endometrium reveal molecular profiles characteristic for specific stages of estrous cycle and early pregnancy. Exp. Clin. Endocrinol. Diabetes 2008, 116, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Bauersachs, S.; Wolf, E. Transcriptome analyses of bovine, porcine and equine endometrium during the pre-implantation phase. Anim. Reprod. Sci. 2012, 134, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Biase, F.H.; Rabel, C.; Guillomot, M.; Hue, I.; Andropolis, K.; Olmstead, C.A.; Oliveira, R.; Wallace, R.; Le Bourhis, D.; Richard, C.; et al. Massive dysregulation of genes involved in cell signaling and placental development in cloned cattle conceptus and maternal endometrium. Proc. Natl. Acad. Sci. USA 2016, 113, 14492–14501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansouri-Attia, N.; Aubert, J.; Reinaud, P.; Giraud-Delville, C.; Taghouti, G.; Galio, L.; Everts, R.E.; Degrelle, S.; Richard, C.; Hue, I.; et al. Gene expression profiles of bovine caruncular and intercaruncular endometrium at implantation. Physiol. Genom. 2009, 39, 14–27. [Google Scholar] [CrossRef] [Green Version]
- McGettigan, P.A.; Browne, J.A.; Carrington, S.D.; Crowe, M.A.; Fair, T.; Forde, N.; Loftus, B.J.; Lohan, A.; Lonergan, P.; Pluta, K.; et al. Fertility and genomics: Comparison of gene expression in contrasting reproductive tissues of female cattle. Reprod. Fertil. Dev. 2016, 28, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Moraes, J.G.N.; Behura, S.K.; Geary, T.W.; Hansen, P.J.; Neibergs, H.L.; Spencer, T.E. Uterine influences on conceptus development in fertility-classified animals. Proc. Natl. Acad. Sci. USA 2018, 115, E1749–E1758. [Google Scholar] [CrossRef] [Green Version]
- Passaro, C.; Tutt, D.; Bages Arnal, S.; Maicas, C.; Laguna-Barraza, R.; Gutierrez-Adan, A.; Browne, J.A.; Rath, D.; Behura, S.K.; Spencer, T.; et al. Global transcriptomic response of bovine endometrium to blastocyst stage embryos. Reproduction 2019. [Google Scholar] [CrossRef]
- Satterfield, M.C.; Song, G.; Kochan, K.J.; Riggs, P.K.; Simmons, R.M.; Elsik, C.G.; Adelson, D.L.; Bazer, F.W.; Zhou, H.; Spencer, T.E. Discovery of candidate genes and pathways in the endometrium regulating ovine blastocyst growth and conceptus elongation. Physiol. Genom. 2009, 39, 85–99. [Google Scholar] [CrossRef] [Green Version]
- Sponchiado, M.; Gomes, N.S.; Fontes, P.K.; Martins, T.; Del Collado, M.; Pastore, A.A.; Pugliesi, G.; Nogueira, M.F.G.; Binelli, M. Pre-hatching embryo-dependent and -independent programming of endometrial function in cattle. PLoS ONE 2017, 12, e0175954. [Google Scholar] [CrossRef]
- Ulbrich, S.E.; Wolf, E.; Bauersachs, S. Hosting the preimplantation embryo: Potentials and limitations of different approaches for analysing embryo-endometrium interactions in cattle. Reprod. Fertil. Dev. 2012, 25, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Elzaiat, M.; Todeschini, A.L.; Caburet, S.; Veitia, R.A. The genetic make-up of ovarian development and function: The focus on the transcription factor FOXL2. Clin. Genet. 2017, 91, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Eozenou, C.; Vitorino Carvalho, A.; Forde, N.; Giraud-Delville, C.; Gall, L.; Lonergan, P.; Auguste, A.; Charpigny, G.; Richard, C.; Pannetier, M.; et al. FOXL2 Is Regulated During the Bovine Estrous Cycle and Its Expression in the Endometrium Is Independent of Conceptus-Derived Interferon Tau. Biol. Reprod. 2012. [Google Scholar] [CrossRef] [PubMed]
- Georges, A.; Auguste, A.; Bessiere, L.; Vanet, A.; Todeschini, A.L.; Veitia, R.A. FOXL2: A central transcription factor of the ovary. J. Mol. Endocrinol. 2014, 52, R17–R33. [Google Scholar] [CrossRef] [PubMed]
- Governini, L.; Carrarelli, P.; Rocha, A.L.; Leo, V.D.; Luddi, A.; Arcuri, F.; Piomboni, P.; Chapron, C.; Bilezikjian, L.M.; Petraglia, F. FOXL2 in human endometrium: Hyperexpressed in endometriosis. Reprod. Sci. 2014, 21, 1249–1255. [Google Scholar] [CrossRef]
- Bellessort, B.; Bachelot, A.; Heude, E.; Alfama, G.; Fontaine, A.; Le Cardinal, M.; Treier, M.; Levi, G. Role of Foxl2 in uterine maturation and function. Hum. Mol. Genet. 2015, 24, 3092–3103. [Google Scholar] [CrossRef]
- Elbaz, M.; Hadas, R.; Bilezikjian, L.M.; Gershon, E. Uterine Foxl2 regulates the adherence of the Trophectoderm cells to the endometrial epithelium. Reprod. Biol. Endocrinol. 2018, 16, 12. [Google Scholar] [CrossRef]
- Al-Gubory, K.H.; Camous, S.; Germain, G.; Bolifraud, P.; Nicole, A.; Ceballos-Picot, I. Reconsideration of the proposed luteotropic and luteoprotective actions of ovine placental lactogen in sheep: In vivo and in vitro studies. J. Endocrinol. 2006, 188, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Krebs, S.; Bauersachs, S.; Blum, H.; Wolf, E.; Miyamoto, A. Actions and interactions of progesterone and estrogen on transcriptome profiles of the bovine endometrium. Physiol. Genom. 2010, 42A, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Crisponi, L.; Deiana, M.; Loi, A.; Chiappe, F.; Uda, M.; Amati, P.; Bisceglia, L.; Zelante, L.; Nagaraja, R.; Porcu, S.; et al. The putative forkhead transcription factor FOXL2 is mutated in blepharophimosis/ptosis/epicanthus inversus syndrome. Nat. Genet. 2001, 27, 159–166. [Google Scholar] [CrossRef]
- Pannetier, M.; Pailhoux, E. Sex differentiation: State of the art and future prospects. Med. Sci. (Paris) 2011, 27, 859–865. [Google Scholar] [CrossRef]
- Uhlenhaut, N.H.; Jakob, S.; Anlag, K.; Eisenberger, T.; Sekido, R.; Kress, J.; Treier, A.-C.; Klugmann, C.; Klasen, C.; Holter, N.I.; et al. Somatic Sex Reprogramming of Adult Ovaries to Testes by FOXL2 Ablation. Cell 2009, 139, 1130–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlenhaut, N.H.; Treier, M. Forkhead transcription factors in ovarian function. Reproduction 2011, 142, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Abdoon, A.S.; Giraud-Delville, C.; Kandil, O.M.; Kerboeuf-Giraud, A.; Eozenou, C.; Carvalho, A.V.; Julian, S.; Sandra, O. Maternal recognition of pregnancy and implantation are not associated with an interferon response of the endometrium to the presence of the conceptus in dromedary camel. Theriogenology 2017, 90, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Bazer, F.W.; Johnson, G.A.; Wu, G. Amino acids and conceptus development during the peri-implantation period of pregnancy. Adv. Exp. Med. Biol. 2015, 843, 23–52. [Google Scholar]
- Franco, H.L.; Jeong, J.W.; Tsai, S.Y.; Lydon, J.P.; DeMayo, F.J. In vivo analysis of progesterone receptor action in the uterus during embryo implantation. Semin. Cell Dev. Biol. 2008, 19, 178–186. [Google Scholar] [CrossRef]
- Lydon, J.P.; DeMayo, F.J.; Funk, C.R.; Mani, S.K.; Hughes, A.R.; Montgomery, C.A., Jr.; Shyamala, G.; Conneely, O.M.; O’Malley, B.W. Mice lacking progesterone receptor exhibit pleiotropic reproductive abnormalities. Genes Dev. 1995, 9, 2266–2278. [Google Scholar] [CrossRef] [Green Version]
- Dinh, D.T.; Breen, J.; Akison, L.K.; DeMayo, F.J.; Brown, H.M.; Robker, R.L.; Russell, D.L. Tissue-specific progesterone receptor-chromatin binding and the regulation of progesterone-dependent gene expression. Sci. Rep. 2019, 9, 11966. [Google Scholar] [CrossRef]
- Lieberman, B.A.; Bona, B.J.; Edwards, D.P.; Nordeen, S.K. The constitution of a progesterone response element. Mol. Endocrinol. 1993, 7, 515–527. [Google Scholar]
- Yin, P.; Roqueiro, D.; Huang, L.; Owen, J.K.; Xie, A.; Navarro, A.; Monsivais, D.; Coon V, J.S.; Kim, J.J.; Dai, Y.; et al. Genome-wide progesterone receptor binding: Cell type-specific and shared mechanisms in T47D breast cancer cells and primary leiomyoma cells. PLoS ONE 2012, 7, e29021. [Google Scholar] [CrossRef] [Green Version]
- Pawar, S.; Hantak, A.M.; Bagchi, I.C.; Bagchi, M.K. Minireview: Steroid-regulated paracrine mechanisms controlling implantation. Mol. Endocrinol. 2014, 28, 1408–1422. [Google Scholar] [CrossRef] [Green Version]
- Goddard, L.M.; Murphy, T.J.; Org, T.; Enciso, J.M.; Hashimoto-Partyka, M.K.; Warren, C.M.; Domigan, C.K.; McDonald, A.I.; He, H.; Sanchez, L.A.; et al. Progesterone receptor in the vascular endothelium triggers physiological uterine permeability preimplantation. Cell 2014, 156, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovici, R.M.; Betzler, N.K.; Krause, M.S.; Luo, M.; Jauckus, J.; Germeyer, A.; Bloethner, S.; Schlotterer, A.; Kumar, R.; Strowitzki, T.; et al. Gene Expression Profiling of Human Endometrial-Trophoblast Interaction in a Coculture Model. Endocrinology 2006, 147, 5662–5675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbi, S.; Hamilton, A.E.; Vo, K.C.; Tulac, S.; Overgaard, M.T.; Dosiou, C.; Le Shay, N.; Nezhat, C.N.; Kempson, R.; Lessey, B.A.; et al. Molecular Phenotyping of Human Endometrium Distinguishes Menstrual Cycle Phases and Underlying Biological Processes in Normo-Ovulatory Women. Endocrinology 2005, 147, 1097–1121. [Google Scholar] [CrossRef] [PubMed]
- Nugent, B.M.; Tobet, S.A.; Lara, H.E.; Lucion, A.B.; Wilson, M.E.; Recabarren, S.E.; Paredes, A.H. Hormonal programming across the lifespan. Horm. Metab. Res. 2012, 44, 577–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pask, A.J.; Calatayud, N.E.; Shaw, G.; Wood, W.M.; Renfree, M.B. Oestrogen blocks the nuclear entry of SOX9 in the developing gonad of a marsupial mammal. BMC Biol. 2010, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pask, A.J. A role for estrogen in somatic cell fate of the mammalian gonad. Chromosome Res. 2012, 20, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Georges, A.; L’Hote, D.; Todeschini, A.L.; Auguste, A.; Legois, B.; Zider, A.; Veitia, R.A. The transcription factor FOXL2 mobilizes estrogen signaling to maintain the identity of ovarian granulosa cells. Elife 2014. [Google Scholar] [CrossRef] [Green Version]
- Pannetier, M.; Fabre, S.; Batista, F.; Kocer, A.; Renault, L.; Jolivet, G.; Mandon-Pepin, B.; Cotinot, C.; Veitia, R.; Pailhoux, E. FOXL2 activates P450 aromatase gene transcription: Towards a better characterization of the early steps of mammalian ovarian development. J. Mol. Endocrinol. 2006, 36, 399–413. [Google Scholar] [CrossRef]
- Govoroun, M.S.; Pannetier, M.; Pailhoux, E.; Cocquet, J.; Brillard, J.P.; Couty, I.; Batellier, F.; Cotinot, C. Isolation of chicken homolog of the FOXL2 gene and comparison of its expression patterns with those of aromatase during ovarian development. Dev. Dyn. 2004, 231, 859–870. [Google Scholar] [CrossRef]
- Baron, D. An evolutionary and functional analysis of FoxL2 in rainbow trout gonad differentiation. J. Mol. Endocrinol. 2004, 33, 705–715. [Google Scholar] [CrossRef]
- Nakamoto, M.; Matsuda, M.; Wang, D.S.; Nagahama, Y.; Shibata, N. Molecular cloning and analysis of gonadal expression of Foxl2 in the medaka, Oryzias latipes. Biochem. Biophys. Res. Commun. 2006, 344, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Georges, A.; Benayoun, B.A.; Marongiu, M.; Dipietromaria, A.; L’Hote, D.; Todeschini, A.L.; Auer, J.; Crisponi, L.; Veitia, R.A. SUMOylation of the Forkhead transcription factor FOXL2 promotes its stabilization/activation through transient recruitment to PML bodies. PLoS ONE 2011, 6, e25463. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Bazer, F.W. Biology of progesterone action during pregnancy recognition and maintenance of pregnancy. Front. Biosci. 2002, 7, d1879–d1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Dunlap, K.A.; Filant, J. Comparative developmental biology of the uterus: Insights into mechanisms and developmental disruption. Mol. Cell. Endocrinol. 2012, 354, 34–53. [Google Scholar] [CrossRef] [PubMed]
- Batista, F.; Vaiman, D.; Dausset, J.; Fellous, M.; Veitia, R.A. Potential targets of FOXL2, a transcription factor involved in craniofacial and follicular development, identified by transcriptomics. Proc. Natl. Acad. Sci. USA 2007, 104, 3330–3335. [Google Scholar] [CrossRef] [Green Version]
- Moumné, L.; Batista, F.; Benayoun, B.A.; Nallathambi, J.; Fellous, M.; Sundaresan, P.; Veitia, R.A. The mutations and potential targets of the forkhead transcription factor FOXL2. Mol. Cell. Endocrinol. 2008, 282, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Geraldo, M.T.; Valente, G.T.; Braz, A.S.; Martins, C. The discovery of Foxl2 paralogs in chondrichthyan, coelacanth and tetrapod genomes reveals an ancient duplication in vertebrates. Heredity (Edinb) 2013, 111, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Wegman, P.; Gothlin Eremo, A.; Lindlof, A.; Karlsson, M.; Stal, O.; Wingren, S. Expression of the forkhead transcription factor FOXL2 correlates with good prognosis in breast cancer patients treated with tamoxifen. Int. J. Oncol. 2011, 38, 1145–1151. [Google Scholar] [CrossRef]
- Sandra, O.; Bataillon, I.; Roux, P.; Martal, J.; Charpigny, G.; Reinaud, P.; Bolifraud, P.; Germain, G.; Al-Gubory, K.H. Suppressor of cytokine signalling (SOCS) genes are expressed in the endometrium and regulated by conceptus signals during early pregnancy in the ewe. J. Mol. Endocrinol. 2005, 34, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Degrelle, S.A.; Campion, E.; Cabau, C.; Piumi, F.; Reinaud, P.; Richard, C.; Renard, J.P.; Hue, I. Molecular evidence for a critical period in mural trophoblast development in bovine blastocysts. Dev. Biol. 2005, 288, 448–460. [Google Scholar] [CrossRef] [Green Version]
- Solano, M.E.; Parker, V.J.; Camous, S.; Sandra, O.; Douglas, A.J.; Arck, P.C. Low doses of trilostane fail to induceabortion during early gestation in miceand ewes. Am. J. Reprod. Immunol. 2008. [Google Scholar] [CrossRef]
- Beard, A.P.; Hunter, M.G.; Lamming, G.E. Quantitative control of oxytocin-induced PGF2 alpha release by progesterone and oestradiol in ewes. J. Reprod. Fertil. 1994, 100, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pant, H.C.; Hopkinson, C.R.; Fitzpatrick, R.J. Concentration of oestradiol, progesterone, luteinizing hormone and follicle-stimulating hormone in the jugular venous plasma of ewes during the oestrous cycle. J. Endocrinol. 1977, 73, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.M.; Healey, G.D.; Sheldon, I.M. Explants of intact endometrium to model bovine innate immunity and inflammation ex vivo. Am. J. Reprod. Immunol. 2012, 67, 526–539. [Google Scholar] [CrossRef]
- Saut, J.P.; Healey, G.D.; Borges, A.M.; Sheldon, I.M. Ovarian steroids do not affect bovine endometrial cytokine or chemokine responses to Escherichia coli or LPS in vitro. Reproduction 2014, 148, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Pannetier, M.; Renault, L.; Jolivet, G.; Cotinot, C.; Pailhoux, E. Ovarian-specific expression of a new gene regulated by the goat PIS region and transcribed by a FOXL2 bidirectional promoter. Genomics 2005, 85, 715–726. [Google Scholar] [CrossRef]
- Kastner, P.; Bocquel, M.T.; Turcotte, B.; Garnier, J.M.; Horwitz, K.B.; Chambon, P.; Gronemeyer, H. Transient expression of human and chicken progesterone receptors does not support alternative translational initiation from a single mRNA as the mechanism generating two receptor isoforms. J. Biol. Chem. 1990, 265, 12163–12167. [Google Scholar]
- Larionov, A.; Krause, A.; Miller, W. A standard curve based method for relative real time PCR data processing. BMC Bioinform. 2005, 6, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eozenou, C.; Lesage-Padilla, A.; Mauffré, V.; Healey, G.D.; Camous, S.; Bolifraud, P.; Giraud-Delville, C.; Vaiman, D.; Shimizu, T.; Miyamoto, A.; et al. FOXL2 is a Progesterone Target Gene in the Endometrium of Ruminants. Int. J. Mol. Sci. 2020, 21, 1478. https://doi.org/10.3390/ijms21041478
Eozenou C, Lesage-Padilla A, Mauffré V, Healey GD, Camous S, Bolifraud P, Giraud-Delville C, Vaiman D, Shimizu T, Miyamoto A, et al. FOXL2 is a Progesterone Target Gene in the Endometrium of Ruminants. International Journal of Molecular Sciences. 2020; 21(4):1478. https://doi.org/10.3390/ijms21041478
Chicago/Turabian StyleEozenou, Caroline, Audrey Lesage-Padilla, Vincent Mauffré, Gareth D. Healey, Sylvaine Camous, Philippe Bolifraud, Corinne Giraud-Delville, Daniel Vaiman, Takashi Shimizu, Akio Miyamoto, and et al. 2020. "FOXL2 is a Progesterone Target Gene in the Endometrium of Ruminants" International Journal of Molecular Sciences 21, no. 4: 1478. https://doi.org/10.3390/ijms21041478